Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Ausubellab.mgh.harvard.edu
Long-Lived C. elegans daf-2
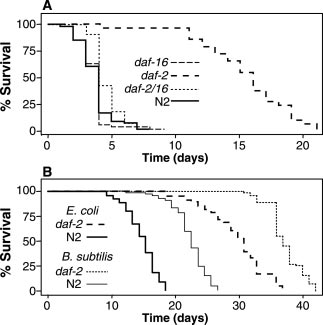
The daf-2 mutant retained extended lon-
gevity relative to wild-type mutants whenpropagated on B. subtilis (Fig. 1B), but the
Mutants Are Resistant to
fractional extension in life-span was modest(76 Ϯ 9%) compared with the fractional ex-
Bacterial Pathogens
tension observed on pathogenic bacteria [E. faecalis, 325 Ϯ 57%; S. aureus, 514 Ϯ189%; P. aeruginosa, 118% Ϯ 14%; and E.
Danielle A. Garsin,1,3 Jacinto M. Villanueva,1,3 Jakob Begun,1,3
coli, 110 Ϯ 11% (9)]. The age-1 mutant also
Dennis H. Kim,1,3 Costi D. Sifri,2,4 Stephen B. Calderwood,2,4
lived longer on B. subtilis (table S1). The
extended survival of the daf-2 and age-1mutants on the relatively innocuous B. subti-lis suggests that additional factors beyond
table S1) under the experimental conditions
pathogen resistance are likely involved in
nisms of aging (1, 2) and immune function
assayed. In these experiments, the daf-2(e1370)
life-span regulation. However, when feeding
(3) in Caenorhabditis elegans. Herein we
allele was more resistant to bacterial pathogens
on pathogenic bacteria, especially Gram-pos-
show that the mechanisms that govern these
than the daf-2(1368) allele and the age-
itive ones, mechanisms underlying pathogen
1(hx546) allele (Fig. 1A; table S1). This is most
resistance appear to be the dominant contrib-
likely a consequence of differences in allele
utor to the overall longevity of daf-2 and
gens Pseudomonas aeruginosa and Salmonella
strengths of the daf-2 and age-1 mutants, all of
enterica and the Gram-positive pathogens En-
which are partial loss-of-function mutants.
Our data suggest that the insulin signaling
terococcus faecalis and Staphylococcus aureus
Because C. elegans daf-2 and age-1 mu-
pathway modulates both inherent longevity
kill C. elegans by an infection-like process with
tants were identified in screens using Esche-
and pathogen resistance to affect overall sur-
remarkable overlap between the bacterial fac-
richia coli strain OP50 as the food source and
vival in a manner dependent on the pathoge-
tors required for virulence in mammals and
because E. coli may also be pathogenic to C.
nicity of the bacteria on which C. elegans is
killing in nematodes (4, 5). Additionally, a p38
elegans (5–7 ), the enhanced longevity phe-
feeding. The linkage of longevity and patho-
notype of daf-2 and age-1 could reflect ac-
gen resistance to the same signaling pathway
signaling cascade is a key component of the
quired resistance to E. coli– mediated killing.
may have general relevance to the observa-
C. elegans innate immune response, as it is
Indeed, nematodes lived considerably longer
tion that most organisms become more sus-
in mammals (3). These experiments estab-
when feeding on the Gram-positive bacteri-
lish C. elegans as a useful model for study-
um Bacillus subtilis, a common soil bacteri-
ing bacterial pathogenicity and host immu-
um that C. elegans is likely to feed on in the
References and Notes
nity. Here we show that certain long-lived
wild (Fig. 1B) (8). It is not likely that B.
1. C. E. Finch, G. Ruvkun, Annu. Rev. Genomics Hum.Genet. 2, 435 (2001). C. elegans mutants are highly resistant to
subtilis is simply more nutritious or more
2. C. A. Wolkow, K. D. Kimura, M.-S. Lee, G. Ruvkun,
readily digestible than E. coli because the rate
Science 290, 147 (2000).
of growth, egg to egg generation time, and
3. D. H. Kim et al., Science 297, 623 (2002).
4. A. Aballay, F. M. Ausubel, Curr. Opin. Microbiol. 5, 97
tween longevity and pathogen resistance, we
number of eggs laid were the same whether
tested whether C. elegans daf-2 and age-1
feeding on B. subtilis or E. coli (fig. S1).
5. D. A. Garsin et al., Proc. Natl. Acad. Sci. U.S.A. 98,
mutants exhibit enhanced resistance to E.
6. D. Garigan et al., Genetics 161, 1101 (2002). faecalis, S. aureus, and P. aeruginosa. daf-2
7. G. V. Mallo et al., Curr. Biol. 12, 1209 (2002).
encodes an insulin-like receptor that functions
8. The difference in life-span between feeding on E. coli
upstream of the phosphatidylinositol 3-kinase
OP50 and B. subtilis PY79 was particularly dramatic
when the longevity assays were carried out at 27°C
(PI 3– kinase) encoded by age-1, and partial loss
rather than at 25°C; the worms lived almost 100%
of function mutations in daf-2 or age-1 result in
longer on B. subtilis then they did on E. coli (table S1).
a long-lived phenotype (1). Both daf-2 and age-1
9. Fractional life-span extension was calculated based
mutants were resistant to killing by E. faecalis, S.aureus, and P. aeruginosa (Fig. 1A; table S1).
10. We thank S. Lee and R. Feinbaum for helpful discussions
Most dramatic was the five- and sixfold in-
and critical reading of the manuscript. D.A.G. and J.M.V.
creased survival of the daf-2(e1370) mutants
are supported by postdoctoral fellowships from the Irv-
ington Institute for Immunological Research. C.D.S. and
relative to wild-type C. elegans when exposed to
D.H.K. are supported by postdoctoral fellowships from
the Gram-positive pathogens E. faecalis and S.
the Howard Hughes Medical Institute. This work was
aureus, respectively (Fig. 1A; table S1).
supported by NIH grant GM48707 (to F.M.A.). Supporting Online Material
www.sciencemag.org/cgi/content/full/300/5627/1921/
insulin-signaling pathway in C. elegans oc-
curs by de-repression of the forkhead tran-
Fig. 1. Pathogen resistance of the C. elegans
scription factor DAF-16, which is normally
daf-2(e1370) mutant. (A) Survival of N2, daf-
under negative regulation by DAF-2. There-
2(e1370), daf-16 (mgDf47), a nd daf-2;daf-16 C.
5 November 2002; accepted 28 February 2003
fore, strong loss-of- function alleles of daf-16elegans feeding on E. faecalis strain OG1RF. (B)
such as mgDf47 suppress the long-lived phe-
Adult life-span of N2 and daf-2(e1370) C. el-
notype of daf-2 mutants (2). daf-16(mgDf47)egans when feeding on B. subtilis (PY79) or E. coli
Department of Genetics, 2Department of Microbiology
(OP50) grown on NG (nematode growth) medi-
and Molecular Genetics, Harvard Medical School, 3De-
also suppressed the pathogen-resistant pheno-
partment of Molecular Biology, 4Division of Infectious
um with FUDR (5-Fluoro-2-deoxyuridine). Assays
type of daf-2(e1370) (Fig. 1A; table S1). Inter-
Diseases, Massachusetts General Hospital, Boston, MA
were carried out as previously described (2, 5).
estingly, the daf-16 mutant exhibited a compa-
STATA 6 statistical software (Stata, College Sta-
rable degree of susceptibility to pathogen-
tion, TX ) was used to plot survival by the Kaplan-
*To whom correspondence should be addressed. E-
mediated killing as wild-type worms (Fig. 1A;
www.sciencemag.org SCIENCE VOL 300 20 JUNE 2003
Hawai`i Department of Agriculture Apiary Program Cause: Paenibacillus larvae , a spore-forming bacterium. Effect: American foulbrood (AFB) is one of the most widespread and the most destructive of the honey bee brood diseases. At first, the population of an infected colony is not no-ticeably decreased and only a few dead larvae or pupae may be present. The disease may develop slowly, wea
PattyLoulookedoutthedoor.Shewaswaitingforhergrandson,Robert,to come. She hadn’t seen him since her ninetieth birthday party threemonths earlier, when the whole family had come out to Brookhaven, Mis-sissippi, to celebrate with her. Robert came up from New Orleans to seeher only three or four times a year, and she was looking forward to seeinghim. She looked out at the sky. There were four buzz
 Long-Lived C. elegans daf-2
Long-Lived C. elegans daf-2