Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
439-450.s9791c-messina
Legumes and soybeans: overview of their nutritional profiles andhealth effects1,2
ABSTRACT
Legumes play an important role in the traditional
countries beans play a less significant dietary role. In fact, bean
diets of many regions throughout the world. In contrast in Western
intake has actually declined during the past century in many
countries beans tend to play only a minor dietary role despite the fact
that they are low in fat and are excellent sources of protein, dietary
In the United States, the availability of dry beans, peas, nuts,
fiber, and a variety of micronutrients and phytochemicals. Soybeans
and soybeans combined has remained fairly constant at 7.3 kg
are unique among the legumes because they are a concentrated
(16 lb), 7.3 kg (16 lb), and 8.2 kg (18 lb) per person per year dur-
source of isoflavones. Isoflavones have weak estrogenic properties
ing the time periods 1909–1913, 1967–1969, and 1985, respec-
and the isoflavone genistein influences signal transduction. Soyfoods
tively (2). For dry edible beans specifically, the annual per capita
and isoflavones have received considerable attention for their poten-
amount available for consumption (product weight) for the years
tial role in preventing and treating cancer and osteoporosis. The low
1972, 1981, 1982, and 1992 was 2.7 kg (6.0 lb), 2.5 kg (5.4 lb),
breast cancer mortality rates in Asian countries and the putative
3.0 kg (6.5 lb), and 3.4 kg (7.5 lb), respectively (3). The 1992
antiestrogenic effects of isoflavones have fueled speculation that
figure represents less than one-quarter servings of beans per per-
soyfood intake reduces breast cancer risk. The available epidemio-
son per day. Less than one-third of the adult US population eats
logic data are limited and only weakly supportive of this hypothe-
beans during any 3-d period (3). The most popular dry bean in
sis, however, particularly for postmenopausal breast cancer. The
the United States is the pinto bean, followed by the navy, kidney,
data suggesting that soy or isoflavones may reduce the risk of
great Northern, and lima bean [annual kg per person for 1995:
prostate cancer are more encouraging. The weak estrogenic effects
1.5 (3.3 lb), 0.8 (1.7 lb), 0.3 (0.6 lb), 0.2 (0.4 lb), and 0.1 (0.2 lb),
of isoflavones and the similarity in chemical structure between soy-
respectively] (4). In the US Department of Agriculture food
bean isoflavones and the synthetic isoflavone ipriflavone, which was
guide pyramid, beans are included in the same group as nuts,
shown to increase bone mineral density in postmenopausal women,
meat, poultry, fish, and seeds (5). Because the recommendation
suggest that soy or isoflavones may reduce the risk of osteoporosis.
is to consume ≥ 2 servings/d from this group, nonvegetarians
Rodent studies tend to support this hypothesis, as do the limited pre-
have relatively little incentive to make beans an important part of
liminary data from humans. Given the nutrient profile and phyto-
chemical contribution of beans, nutritionists should make a con-
Beans tend to have a poor image and one that stands in stark
certed effort to encourage the public to consume more beans in
contrast to the nutritional value they offer. Beans have been called
general and more soyfoods in particular.
the “poor man’s meat,” a metaphor which is consistent with the
inverse relation between bean intake and income. For US malesaged ≥ 20 y, the frequency of bean intake during a 3-d period was
KEY WORDS
Legumes, soybeans, beans, phytochemicals,
36.3%, 32.3%, and 25.7% among men with incomes < 131%,
isoflavones, genistein, soyfoods, breast cancer, prostate cancer,
131–350%, and > 350% of the poverty level, respectively (3).
cancer prevention, osteoporosis prevention, dietary fiber
Given the important role of beans in populations that consume
plant-based diets, it is not surprising that legume intake is higher invegetarians than in nonvegetarians, although the data are limited (6,
INTRODUCTION
7). Certainly, one would expect the consumption of beans to
Legumes include peas, beans, lentils, peanuts, and other pod-
increase with the elimination of meat and eggs from the diet by lac-
ded plants that are used as food. Legumes have been cultivated
tovegetarians and vegans. Appropriately, the vegetarian food guide
for thousands of years, although many of the varieties of beans
pyramid recently developed by Loma Linda University places
and peas that are commonplace today were unknown until rela-
legumes in their own group at the bottom of the pyramid (8).
Legumes have played an important role in the traditional diets
of many regions throughout the world. It is difficult to think ofthe cuisines of Asia, India, South America, the Middle East, and
1 From Nutrition Matters, Inc, Townsend, WA.
Mexico without picturing soybeans, lentils, black beans, chick-
2 Address reprint requests to MJ Messina, Nutrition Matters, Inc, 1543
peas, and pinto beans, respectively. In contrast, in many Western
Lincoln Street, Port Townsend, WA 98368. E-mail: markm@olympus.net. Am J Clin Nutr 1999;70(suppl):439S–50S. Printed in USA. 1999 American Society for Clinical Nutrition
Beans have long been recognized for their protein content and
value for protein quality. The PDCAASs of most beans are rea-
more recently have been noted for their soluble-fiber content, but
sonably good, although their overall value is reduced somewhat
in general there has been relatively little research and discussion
by their lower digestibility (19). Some types of soy protein prod-
about the nutritional attributes of legumes. The glaring exception
ucts have PDCAASs of close to one, the highest value possible.
to this is the soybean, which has been investigated intensively dur-
Some concerns have been raised about the use of the PDCAAS
ing the past 5–10 y. This is largely because soybeans are a unique
(20), but it certainly represents an improvement over the protein-
dietary source of a group of phytochemicals called isoflavones.
Isoflavones are thought to exert a myriad of biological effects and
Ironically, the relatively low SAA content of beans may actu-
it has been hypothesized that they reduce the risk of a number of
ally provide an advantage in terms of calcium retention. The
reported hypercalciuric effect of protein is likely to be at least par-
This article provides an overview of the nutritional attributes
tially due to the metabolism of SAAs. The skeletal system serves
of dry beans in general, and then focuses on soybeans in relation
as one of the main buffering systems in the body; as a result, the
to risk of breast and prostate cancers and osteoporosis. The
hydrogen ions produced from the metabolism of SAAs cause dem-
reader is referred to other articles in this supplement for reviews
ineralization of bone and excretion of calcium in the urine (21,
on nuts (9, 10), additional information on legumes (11), and a
22). Thus, bean protein may improve calcium retention relative to
discussion of the effects of soy in relation to heart and kidney
animal and grain proteins. In general, it has been estimated that
every gram of protein consumed causes the loss of 1 mg Ca (23). Although this may appear to be a trivial amount, every additionalmilligram of calcium excreted may markedly increase dietary cal-
NUTRIENT COMPOSITION
cium requirements because net calcium absorption is substantiallyless than the average calcium absorption (Ϸ30%) from foods.
Human studies showed that the consumption of soy protein is
The macronutrient composition of selected beans is shown in
associated with a markedly lower urinary calcium excretion com-
Table 1. The protein content of beans is generally between 20%
pared with the consumption of similar amounts of whey protein
and 30% of energy. A serving of beans (Ϸ90 g or 1/2 cup cooked
(24) or a mixture of animal proteins (25).
beans) provides Ϸ7–8 g protein or Ϸ15% of the recommended
With regard to bone health, the nutritional significance of sub-
dietary allowance (RDA) for protein for a 70-kg adult (15).
stituting bean protein for animal protein depends on the relative
Although legumes are recognized as being high in protein, the
amounts consumed. In general, this process would appear to play
quality of bean protein is often underestimated. This is because
a minor role because legume protein, even among populations
the protein-efficiency ratio, which is based on the growth of lab-
eating plant-based diets, comprises only a small percentage of
oratory animals (most commonly rats), was the standard method
total protein intake. However, the hypocalciuric effect of bean
of evaluating protein quality until recently. Rats have a methion-
proteins may be quite important for some individuals, such as
ine requirement that is Ϸ50% higher than that of humans (16).
those substituting soy protein for animal protein because of its
Consequently, because bean proteins are relatively low in sulfur
reported hypocholesterolemic effect (12) and athletes using soy
amino acids (SAAs), the protein-efficiency ratios of beans are
protein supplements. It should be noted, however, that not all
studies are in agreement about the effects of protein on calcium
However, the World Health Organization (WHO) and the US
Food and Drug Administration have adopted an alternativemethod for evaluating protein quality called the protein digest-
ibility corrected amino acid score (PDCAAS) (18). This method
Most beans are very low in fat, generally containing Ϸ5% of
uses the amino acid score (based on the Food and Agriculture
energy as fat (Table 1). The primary exceptions are chickpeas and
Organization estimated amino acid requirement for 2–5-y-old
soybeans, which contain Ϸ15% and 47% fat, respectively. The
children) and a correction factor for digestibility to arrive at a
predominant fatty acid in beans is linoleic acid, although beans
TABLE 1 Nutrient content of selected beans (serving size is Ϸ90 g or 1⁄ c boiled)1 1 From reference 13. 2 Value represents crude fiber. From reference 14.
also contain the nϪ3 fatty acid, ␣-linolenic acid (28). However,
index were Ϸ40% more likely to develop diabetes than those
because the overall fat content of most beans is so low, the dietary
consuming low-glycemic-index diets, even after controlling for
contribution of beans to ␣-linolenic acid intake is generally minor.
several diabetes risk factors (55). Thus, beans may be a particu-
As noted, soybeans are quite high in fat, and the consumption of
larly important food for individuals with diabetes and those with
full-fat soyfoods contributes significantly to ␣-linolenic acid
an elevated risk of developing diabetes.
intake. The ratio of linoleic to ␣-linolenic acid in soybeans is
Ϸ7.5:1 (␣-linolenic acid makes up Ϸ7–8% of the total fat) (28). Nonnutritive components
nϪ3 Polyunsaturated fatty acids, especially eicosapentaenoic acid
Beans contain several components that traditionally have been
(EPA) and docosahexaenoic acid (DHA), are being studied for
considered to be antinutrients, such as trypsin inhibitors, phytate
their health benefits (29–32). Adequate DHA status is particularly
(inositol hexaphosphate), oligosaccharides, and saponins. More
important for infants (33). ␣-Linolenic acid can be converted into
recent information suggests, however, that the antinutrient label
EPA and EPA can be converted into DHA, although the rate of
may be an oversimplification, especially in the case of oligosac-
conversion of ␣-linolenic acid into EPA is relatively inefficient, at
charides and saponins. Trypsin inhibitors from beans can cer-
5–10% (34, 35), and is inhibited by linoleic acid (34). The dietary
tainly interfere with protein digestion, and in some species of
ratio of nϪ6 to nϪ3 fatty acids among vegetarians (36) is at the
animals do cause pancreatic enlargement and enhance chemi-
high end of the rather conservative recommendations by the WHO
cally induced pancreatic tumors (56). However, boiling dry
(5:1–10:1) (37). The nϪ3 status of vegetarians is an issue that
beans generally reduces the trypsin inhibitor content by 80–90%
(57) and there is little reason to think that the amount of trypsininhibitors obtained by eating commonly consumed beans would
Micronutrients
exert any adverse effects in humans (58). In contrast to the
The folate, iron, zinc, and calcium contents of selected beans
trypsin inhibitor, the trypsin and chymotrypsin inhibitor (Bow-
are listed in Table 1. Beans are an excellent source of folate, which
man-Birk inhibitor) found in beans, especially soybeans, has
in addition to being an essential nutrient is thought to reduce the
been studied as an anticancer agent (59).
risk of neural tube defects (38). One serving of beans provides
As noted above, phytate is thought to contribute to the poor
more than half of the current RDA for folate (15). Beans are also
mineral bioavailability of beans. On average, the phytate con-
high in iron; 1 serving provides Ϸ2 mg. This compares favorably
centration in beans is between 1% and 2% (60, 61). Although the
with the iron RDAs of 10 and 15 mg for adult men and pre-
effect of phytate in reducing mineral bioavailability in plant
menopausal women, respectively (15). However, iron bioavail-
foods is an important consideration, it has also been postulated
ability from legumes is poor and thus their value as a source of
that phytic acid may play a role in reducing cancer risk, possibly
iron is diminished (39). In acute studies, the addition of vitamin C
because of its antioxidant effects (62). Specifically, it has been
to foods markedly increased nonheme iron absorption (40), but in
suggested that phytic acid may lower the risk of colon cancer
longer-term studies the effects of vitamin C intake on iron absorp-
(63) and perhaps breast cancer (64).
tion and status were much less pronounced (41). In general, sin-
More than 40 y ago, diets containing beans were first shown
gle-meal studies overestimate the effects of both inhibitors and
to markedly increase flatulence (65). In 1970, it was reported
enhancers of nonheme iron absorption (42).
that the oligosaccharides in beans were responsible for gas pro-
In contrast to iron bioavailability, zinc bioavailability from
duction (66). The oligosaccharide content of dry beans is
legumes is relatively good at Ϸ25% (43). Also, many beans are
Ϸ25–50 mg/g (67, 68). Because there is no ␣-galactosidase in
good sources of calcium, providing on average Ϸ50 mg Ca/serv-
the human intestinal mucosa to cleave the ␣-(1–6) galactose
ing, although there is quite a bit of variation among the legumes.
linkage present in galactoside-containing oligosaccharides, such
Calcium bioavailability from beans in general is Ϸ20%, which is
as raffinose and stachyose, these oligosaccharides pass into the
lower than that from milk and green leafy vegetables but is still
large intestine where bacteria metabolize them and form large
reasonably good (44). Calcium bioavailability from soybeans
amounts of carbon dioxide, hydrogen, and sometimes methane.
and soyfoods is quite good—essentially equivalent to calcium
Because of the discomfort and social embarrassment associated
bioavailability from milk—despite the fact that soybeans are
with flatulence, some people opt to avoid beans entirely.
Commercial products such as Beano (AkPharma Inc, Pleas-
antville, NJ), a digestive aid that contains ␣-galactosidase, are
Fiber and the glycemic index
available so that individuals can eat beans without discomfort.
Beans are an excellent source of dietary fiber; 1 serving pro-
Additionally, it is possible to remove substantial amounts of
vides 2–4 g of a mix of soluble and insoluble fiber (46). High-
oligosaccharides and to markedly reduce flatulence by changing
fiber, high-bean diets were shown to lower serum cholesterol in
the water in which beans are boiled one or more times (69).
hypercholesterolemic individuals (47). In addition, beans have
However, there may be some beneficial effects associated with
very low glycemic indexes (48, 49). This has been attributed to
oligosaccharide consumption. The oligosaccharides, because of
many factors including their fiber (50), tannin (51), and phytic
their growth-promoting effect on bifidobacteria, have been
acid contents (52). Although neither the American Diabetes
hypothesized to promote the health of the colon, increase
Association nor the American Dietetic Association endorse the
longevity, and decrease colon cancer risk (70–72). In fact, for
glycemic index as a tool for constructing diets for individuals
these reasons researchers in Japan have actually suggested that
with diabetes (53), research published during the past decade
soybean oligosaccharides be used as a substitute for common
makes a persuasive argument that the glycemic index of foods is
table sugar (73). For a more detailed discussion of oligosaccha-
one factor affecting the overall quality of the diet (54). In sup-
rides, see Slavin et al in this supplement (74).
port of this statement are findings from a prospective study
Saponins are glycosides composed of a lipid-soluble aglycone
showing that women who consumed diets with a high glycemic
that consists of either a sterol or, more commonly, a triterpenoid
structure attached to water-soluble sugar residues that differ in
as the conjugate, whereas in fermented soy products such as
their type and amount. The major sources of dietary saponins are
legumes, and many types of saponins can be present in the same
In addition to the isoflavones found in soybeans, the intestinal
bean. Saponins are very poorly absorbed. Most saponins form
microflora can convert daidzein into several different products,
insoluble complexes with 3--hydroxysteroids and are known to
including the isoflavonoids equol (7-hydroxyisoflavan), dihydro-
interact with and form large, mixed micelles with bile acids and
daidzein, and O-desmethylangolensin (80). However, because of
cholesterol. Although saponins were shown to lower cholesterol in
differences in intestinal microflora, equol production occurs in
some animal species, the hypocholesterolemic effects of saponins
only Ϸ1 out of every 3 individuals consuming soyfoods (81, 82).
in humans are more speculative (75). Saponins may have anti-
It has been proposed that in humans, genistein is metabolized to
cancer properties, as suggested by a recent rodent study that found
dihydrogenistein and 6Ј-hydroxy-O-desmethylangolensin (80).
that a saponin-containing diet (3% by wt) inhibited by about two-
Estrogenic and antiestrogenic activity
thirds the development of azoxymethane-induced preneoplasticlesions in the colon (76). However, given that human intake of
Initial interest in the beneficial effects of isoflavones focused
saponins is generally ≤ 200–300 mg/d whereas total food intake is
on their estrogenic activity and their possible use in the animal
Ϸ500 g (dry weight), it is not clear to what extent these results in
feed industry as growth promoters (83). On a molar basis relative
to physiologic estrogens, isoflavones are quite weak according to
Isoflavones make up another group of phytochemicals found
both in vitro and in vivo assays, possessing between 1 ϫ 10–4
in beans, but for practical purposes the soybean is the only nutri-
and 1 ϫ 10–3 the activity of 17-estradiol (84–90). Despite their
tionally relevant source of these compounds. Soybeans and soy
relatively low potency, isoflavones are likely to exert physiologic
products contain Ϸ1–3 mg isoflavones/g protein; 1 serving of
effects because it has been shown that in people who consume
traditional soyfoods provides Ϸ25–40 mg isoflavones (77, 78).
soyfoods, serum concentrations of isoflavones are several orders
Isoflavones have received considerable attention in recent years.
of magnitude higher than those of physiologic estrogens. Studies
They are being studied for their potential role in the prevention
have found that, in response to the consumption of soyfoods,
and treatment of a number of chronic diseases including certain
blood isoflavone concentrations can reach the low micromolar
forms of cancer, osteoporosis, and heart disease, and also for
range (≤ 6 mol/L) (91), although concentrations in free-living
their ability to relieve menopausal symptoms.
Japanese men are generally in the high nanomolar range (300–400 nmol/L) (92). Soybean isoflavones
Although isoflavones are weak estrogens, Folman and Pope
Isoflavones are a subclass of the more ubiquitous flavonoids.
(84) showed > 30 y ago that in female mice genistein injected
The basic structural feature of flavonoid compounds is the
subcutaneously inhibited estrone stimulation of uterine growth;
flavone nucleus, which is composed of 2 benzene rings (A and
thus, the authors concluded that genistein could function as an
B) linked through a heterocyclic pyrane C ring (Figure 1). The
antiestrogen. The prevailing hypothesis has been that isoflavones
position of the benzenoid B ring is the basis for dividing the
exert antiestrogenic effects when placed in a high-estrogen envi-
flavonoid class into flavonoids (2-position) and isoflavonoids
ronment, such as exists in premenopausal women, and estrogenic
(3-position). The primary isoflavones in soybeans are genistein
effects when in a low-estrogen environment, such as exists in
(4Ј,5,7-trihydroxyisoflavone) and daidzein (4Ј,7-dihydroxy-
postmenopausal women. This hypothesis has some support; for
isoflavone) and their respective -glycosides, genistin and
example, Mäkela et al (93) found that in ovariectomized mice
daidzin (sugars are attached at the 7 position of the A ring).
not given the synthetic estrogen diethylstilbestrol (DES), uterine
Much lower amounts of glycitein (7,4Ј-dihydroxy-6-methoxy-
weight increased in those fed soy compared with control animals
isoflavone) and its glycoside, glycitin, are present in soybeans
(0.87 and 0.76 mg/g body wt, respectively; P < 0.001). In mice
(79). In nonfermented soyfoods, the isoflavones appear mostly
given DES, uterine weight decreased in those fed soy comparedwith control animals (1.01 and 1.49 mg/g body wt, respectively;P < 0.001).
In addition to competing with endogenous estrogens for bind-
ing to the estrogen receptor, there are several potential mecha-nisms by which the isoflavones may exert antiestrogenic effects(reviewed in 94). However, there are conflicting results aboutwhen isoflavones and soy exert hormonal effects and whetherthese effects are estrogenic or antiestrogenic in nature (95–106). This should not be surprising given recent insights into the intri-cacy of the ligand-estrogen receptor binding complex (reviewed in107) and the identification of a novel, second estrogen receptor, ,to which isoflavones bind (108). Particularly germane to this issue,however, are the findings of 2 human studies suggesting that soyconsumption exerts estrogenic effects on breast tissue. Epidemio-logic research by Wrensch et al (109) showed that breast-nipple-aspirate fluid is a biomarker for breast cancer risk. Women whosecrete fluid are at increased risk compared with nonsecretors, andwomen who secrete fluid containing cells with abnormal cytology(eg, hyperplastic cells) are also at increased risk. In a 9-mo study
FIGURE 1. Structures of the primary isoflavones in soybeans.
by this group, contrary to expectations, breast fluid secretion in
both premenopausal and postmenopausal women taking hormone
addition to the low breast cancer mortality rates in Asia, 2 other
replacement therapy increased in response to soy consumption, as
early observations provided a basis for the hypothesis that soy
did the number of atypical cells in the breast fluid (110). However,
intake decreases breast cancer risk: 1) the potential antiestrogenic
this was a pilot study that did not include a control group.
effects of the soybean isoflavones as discussed above, and 2) the
In a recent study of premenopausal women by McMichael-
reduced number of 7,12-dimethylbenz(a)anthracene–induced
Phillips et al (111), the rate of DNA synthesis by breast cells
mammary tumors observed in rats fed a diet containing soy (142).
taken from biopsies of normal breast tissue from women with
Since this hypothesis was initially proposed, several epidemio-
benign or malignant breast disease was enhanced by 2 wk of soy
logic studies have examined the relation between soy intake and
feeding. Although the clinical implications of this study and the
study by Petrakis et al (110) are a matter of debate, when the in
In 1991, a case-control study conducted by Lee et al (143) in
vitro, animal, and human data are considered it is difficult to
Singapore (n = 200 case subjects and 420 control subjects) found
conclude that soy or isoflavones are necessarily antiestrogenic in
that regular consumption of soyfoods was associated with a
marked decrease in breast cancer risk in premenopausal women(odds ratio: 0.39; 95% CI for the highest fifth compared with the
Effects of soy and isoflavones on cancer risk
lowest fifth of intake of total soy products: 0.19, 0.77; P < 0.02)
Data regarding the relation between soy intake and cancer risk,
but not postmenopausal women. A Japanese case-control study
including in vitro, animal, and epidemiologic results, were
(n = 1186 case subjects and 23 163 control subjects) also found
reviewed by Messina et al (112). On the basis of this review, it is
that tofu intake (≥ 3 times/wk compared with < 3 times/wk) was
clear that the data are insufficient to conclude that soy consump-
associated with decreased risk of breast cancer in premenopausal
tion is protective, and yet the data certainly warrant continued
women (odds ratio: 0.81; 95% CI: 0.65, 0.99; P < 0.05), but
investigation of this relation. Besides isoflavones, there are a num-
again, soy intake was not protective against postmenopausal
ber of phytochemicals in soybeans with demonstrated anticarcino-
breast cancer (144). In contrast to these studies, a case-control
genic activity; these include phytosterols, phytates, saponins, pro-
study involving 2 different locations in China [Shanghai (n = 534
tease inhibitors, and a variety of phenolic acids (113). However,
case subjects and 534 control subjects) and Tianjin (n = 300 case
most of the data point toward the isoflavones as being responsible
subjects and 300 control subjects)] failed to find an association
for the hypothesized anticancer effects of soy.
between soyfood intake and breast cancer risk in either pre- or
Daidzein, one of the 2 primary isoflavones in soybeans,
exhibits anticancer effects; eg, it inhibited the growth of HL-60
The only case-control study (n = 596 case subjects and 958
cells implanted in the subrenal capsules of mice (114). However,
control subjects) conducted thus far in the United States to exam-
genistein has attracted most of the interest. There are literally
ine the relation between soy intake and breast cancer risk found
hundreds of in vitro studies showing that genistein inhibits the
that tofu consumption was protective in both premenopausal
growth of a wide range of both hormone-dependent and hormone-
(adjusted odds ratio: 0.67) and postmenopausal (adjusted odds
ratio: 0.70) Asian women (146). However, the overall intake of
(2–10 g/mL), including breast (115–121), prostate (122–124),
tofu among the subjects in this study was relatively low; the high-
colon (125, 126), and skin (127) cells (reviewed in 94, 128, 129).
est quartile of intake included women who consumed tofu as
Also, in vitro, genistein inhibits the metastatic activity of both
infrequently as 55 times per year. Also, the protective effect was
breast (130) and prostate (131) cancer cells independent of the
primarily in Asian women born in Asia who migrated to the West
and not in Asian Americans born in the United States (146). One
Although the antioxidant properties of genistein may con-
interpretation of these findings is that tofu intake per se is not pro-
tribute to the anticancer effects observed in vitro (132), it is more
tective but rather that it is simply reflective of some protective
likely that these effects are due to the inhibitory actions of genis-
lifestyle common to women of Asian ancestry born in Asia but
tein on several enzymes involved in signal transduction, includ-
not those born in the United States. Alternatively, the anticancer
ing tyrosine protein kinases (133), MAP kinase (134), and riboso-
effects of tofu may be negated by a lifestyle common to women
mal S6 kinase (135). Genistein also inhibits the activity of DNA
of Asian ancestry born in the United States but not those born in
topoisomerase II (136) and Peterson et al (137) recently reported
Asia. Finally, in the Iowa Women’s Study, a prospective study
that genistein increased the in vitro concentrations of transforming
involving > 34 000 women, it was found after 8 y of follow-up
growth factor  (TGF). This last finding may be particularly
that tofu intake was associated with a modest decrease in post-
important given the role that TGF may have in inhibiting the
menopausal breast cancer risk (adjusted relative risk for any con-
growth of cancer cells (138–140). Although there are in vitro, ani-
sumption compared with no consumption: 0.76), although this
mal, and epidemiologic data supporting a protective role of soy or
was not a statistically significant effect (P < 0.22) (147). Not
isoflavones against several forms of cancer, this review will con-
unexpectedly, only 2.9% of the cohort reported eating any tofu.
sider only breast and prostate cancers because most of the focus
Overall, the epidemiologic data are inconclusive. There is rel-
atively little epidemiologic support for the notion that soy intakeis associated with a decreased risk of postmenopausal breast can-
cer. However, there are some limited data, albeit inconsistent,
Research on the relation between soy intake and cancer risk ini-
suggesting that soy intake is associated with a decreased risk of
tially focused primarily on cancer of the breast. In large part, inter-
est in this relation was due to the relatively low breast cancer mor-
As noted previously, genistein has been shown to inhibit the
tality rates in Asian countries where soyfoods are commonly
growth of both estrogen-dependent and estrogen-independent
consumed. In Japan for example, the breast cancer mortality rate
breast cancer cells in vitro, but it is not clear that cellular con-
is only about one-quarter of that of the United States (141). In
centrations of genistein in vivo would reach the in vitro concen-
trations required to inhibit breast cancer–cell growth. It should
soyfoods may be a factor contributing to the low prostate cancer
be noted, however, that Peterson and Barnes (148) found that
mortality rate in Japan, although the data in support of this
genistein inhibits the serum and epidermal growth factor–stimu-
hypothesis, while intriguing, are limited.
lated growth of normal human mammary epithelial cells with
Genistein inhibits the growth of both androgen-dependent and
values 11–15-fold lower than those for human transformed
androgen-independent prostate cancer cells in vitro (122, 123).
breast epithelial cells. Thus, soy intake may help to prevent the
Genistein also inhibits the metastatic potential of prostate cancer
initiation of cancer cells, rather than inhibiting the growth of
cells independent of cell growth inhibition, an effect that is asso-
ciated with a decrease in the tyrosine phosphorylation of an
In a study by Constantinou et al (149), neither genistein nor
unidentified molecular species (131).
daidzein (injected intraperitoneally) inhibited N-methyl-N-
In addition to the effects of genistein on signal transduction
nitrosourea–induced mammary tumor incidence in Sprague-
that were noted previously, there are other mechanisms by which
Dawley rats, although both isoflavones had a moderate but not
genistein or isoflavones could reduce prostate cancer risk. For
statistically significant effect on tumor multiplicity (6.7 com-
example, even though the precise role of estrogen in prostate can-
pared with 4.9 tumors/rat). Because synergistic effects between
cer is not well defined, the potential estrogenic effects of iso-
genistein and daidzein have been noted in vitro, it would be of
flavones may be protective because estrogens have been used suc-
interest to examine their combined effects in vivo (150, 151). Of
cessfully as a form of hormone therapy for metastatic prostate
course there is also the possibility that other components of soy-
cancer (156). Also, some data indicate that genistein inhibits the
beans, individually or in conjunction with isoflavones, are
activity of 5-␣-reductase in genital skin fibroblasts and benign
responsible for the hypothesized anticancer effects of soyfoods.
hyperplastic prostate tissue (150). This enzyme converts testos-
It is apparent from the human studies by Wrensch et al (109),
terone into the more active form of androgen, dihydrotestos-
McMichael-Phillips et al (111), and Cassidy et al (103, 104) that
terone, which stimulates the growth of prostate tissue. Ross et al
soy or isoflavones have the potential to exert physiologic effects
(157) showed that biomarkers of 5-␣-reductase activity are higher
theoretically related to breast cancer risk. In particular, Cassidy
in white and black men compared with Japanese men. The in
et al (103) found that the consumption of soy, specifically
vitro data from Evans et al (150) are consistent with findings from
isoflavone-rich soy (104), extends the length of the follicular
Lu et al (158), who reported that after 1 mo of soymilk consump-
phase and decreases serum follicle-stimulating hormone and
tion (36 oz/d), serum concentrations of 3␣,17-androstanediol
luteinizing hormone concentrations. It is certainly not possible to
glucuronide, a metabolite of dihydrotestosterone, were signifi-
conclude at this time that consumption of soyfoods in adulthood
is a factor that contributes to the low breast cancer mortality
Until recently, there were few animal studies related to soy and
rates among Japanese and Asian women, although this hypothe-
prostate cancer. In 1995, Mäkela et al (93) reported that after
sis still warrants rigorous investigation.
feeding mice a diet containing soy for 9 mo, the incidence of pro-
Finally, there are provocative data from Brown and Lamar-
static dysplasia, which may viewed as a preneoplastic prostate
tiniere (152), Lamartiniere et al (153), and Murrill et al (154) sug-
lesion, was markedly reduced compared with the incidence in
gesting that the early consumption of soyfoods by young girls
mice fed a diet not containing soy (30% and 80%, respectively).
may reduce breast cancer development later in life. This research
At 12 mo, however, there was much less difference between the
group has shown that early exposure (during the neonatal or pre-
2 groups (64% compared with 86%). These findings are consis-
pubertal period of life) to genistein (subcutaneous administration)
tent with the epidemiologic data noted above and also with the
inhibits the development of dimethylbenz(a)anthracene-induced
results of a study of MNU-induced prostate tumors in Lobund-
mammary tumors in rodents and increases the latency period
Wistar rats (159). Rats fed a diet containing soy with a low
(152–154). These findings offer a potential explanation for the
amount of isoflavones had a shorter latency period [7.3 mo for
findings of Wu et al (146). Perhaps Asian women born in Asia are
pre-MNU group and 9.3 mo for post-MNU group] than those fed
exposed to tofu at an earlier age than Asians born in the West.
a diet containing soy high in isoflavones [10 mo for pre-MNU
Certainly, the work of this group provides the basis for an intrigu-
group and 10.6 mo for post-MNU group] (159).
ing line of investigation, especially because recent research indi-
Three studies examined the effect of soy or genistein on tumor
cates that early dietary exposure to genistein is also effective in
development in rats implanted with prostate cancer cells (123,
retarding later development of mammary cancer.
160, 161). Zhang et al (160) found that in rats fed a diet contain-ing soy flour (33% by weight) and implanted with Dunning R3327
PAP tumors, tumor growth was significantly retarded at 16 wk
As is the case for breast cancer, prostate cancer mortality rates
compared with animals fed the control diet. Schleicher et al (161)
vary markedly among countries. An interesting observation
found that genistein (50 mg/kg body wt) given under the skin in
related to the occurrence of prostate cancer is that rates of clini-
the dorsal scapular area every 12 h starting at the time of tumor
cal prostate cancer vary much more than rates of latent prostate
cell transplantation inhibited the development of prostate tumors
cancer. For example, the US incidence of clinical prostate cancer
in rats implanted with prostate carcinoma cells. Rats given genis-
among whites is 10–15-fold higher than the Japanese rate,
tein developed fewer tumors and fewer invasive tumors, and no
whereas the overall incidence of latent prostate cancer is only
genistein-treated animals developed lung metastases.
Ϸ50% higher (155). This suggests that in some populations,
Insight into a possible mechanism for the inhibitory effects of
such as the Japanese, the growth of prostate tumors is slower, the
genistein came from Dalu et al (162), who found that in Lobund-
onset of prostate tumors occurs later in life, or both. Delaying the
Wistar rats, dietary genistein (1 mg genistein/g diet) reduced the
appearance of clinical prostate tumors by even a few years could
weight of the dorsolateral and ventral prostates and inhibited the
have a marked impact on mortality because prostate cancer typ-
expression of tyrosine-phosphorylated proteins. This study was
ically occurs in older men. There is speculation that the intake of
the first to show that in vivo, genistein inhibits a key cellular path-
way. Related to this finding are those of Geller et al (163), who
larger, follow-up study is currently underway. Interestingly, the
found that genistein (at concentrations of 1–15 g/mL) inhibited
genes in which hereditary hemorrhagic telangiectasia mutations
the incorporation of 3H-thymidine (a measure of tissue growth)
have been mapped thus far all encode for proteins that are involved
in cultured benign prostatic hypertrophy tissue by 44–86% in a
in TGF signaling (169) and as noted previously, Peterson et al
showed that in vitro, genistein increases TGF levels (137).
In contrast to the favorable results discussed above, Naik et al
(123) found that although genistein inhibited prostate cancer cell
Soy and bone health
growth in vitro, when Copenhagen rats were injected in the right
The similarity in structure between the isoflavones and estro-
flank with the metastatic MAT-Lylu prostate cancer line, oral
gen and the findings that isoflavones possess weak estrogenic
doses of genistein (0.07, 0.143, and 0.285 mg/d) failed to inhibit
properties as shown by various experimental models provided the
the development of prostate tumors. These doses more closely
initial basis for speculation that isoflavones may promote bone
approximated human dietary intake than the amounts used by
health. Speculation about the potential benefits of isoflavones
Schleicher et al (161) and Dalu et al (162). Higher doses of genis-
was also fueled by the similarity in chemical structure between
tein (0.143, 0.285, and 0.428 mg/kg) injected by the intraperi-
the soybean isoflavones and the synthetic isoflavone, 7-isoprop-
toneal route also had little effect on tumor growth (123).
oxyisoflavone (ipriflavone), which was shown to increase bone
Not surprisingly, there are limited human data available for use
mass in postmenopausal women (170, 171).
in evaluating the soy–prostate cancer hypothesis, although a
Interestingly, for ipriflavone to be maximally effective it
prospective study by Severson et al (164) found that consumption
requires metabolism, and one of the metabolites of ipriflavone is
of tofu was associated with a markedly reduced risk of prostate
the soybean isoflavone daidzein (166). The usual dose of ipri-
cancer (age-adjusted relative risk: 0.35 for subjects who ate tofu
flavone is between 600 and 1200 mg/d. Reportedly, daidzein
≥5 times/wk compared with those who ate tofu ≤1 time/wk).
comprises 10% of the metabolic products of ipriflavone (171),
However, this difference did not quite reach statistical significance
although it is not clear to what extent daidzein is actually respon-
(P < 0.054) and the number of men with tumors in each of the ter-
sible for the effects of ipriflavone on bone resorption; it appears
tiles was small (164). Of potential relevance to the effects of
to be one of several metabolites able to inhibit osteoclast activity
isoflavones on prostate cancer risk is the finding that isoflavones
appear in the prostatic fluid, and that concentrations are highest in
The lower rate of hip fracture among Japanese women in com-
men from soyfood-consuming countries (165). Furthermore, rela-
parison to US women (173, 174) is often cited as providing sup-
tive to plasma concentrations, isoflavones are concentrated sev-
port for a protective effect of isoflavones, but this line of reason-
eral-fold in the prostatic fluid. Interestingly, a recent case study
ing appears to be without merit. The bone density of Japanese
reported significant apoptosis in a prostatic specimen from a man
women is similar to or lower than that of US women, whose hip
with adenocarcinoma who had taken isoflavones (160 mg/d)
fracture rate is twice as high (175–177). Furthermore, the Japan-
derived from red clover 1 wk before surgery. The red clover
ese vertebral fracture rate is actually much higher than that of US
extract contains both genistein and daidzein as well as the methy-
women (176). The low Japanese hip fracture rate is thought to be
lated isoflavones, biochanin-A and formononetin, from which
due at least in part to anatomical differences between white and
genistein and daidzein, respectively, are derived (166).
Japanese women, such as the shorter hip axis length of Japanesewomen (178), and perhaps also to other factors such as a lower
There has been some speculation that soy or isoflavones could
Until recently there were no direct data indicating that the soy-
be used in the treatment of existing tumors, either alone or in con-
bean isoflavones affect bone density. In 1995, Anderson et al (180)
junction with conventional chemotherapeutic agents. Support for
reported that genistein exhibited a biphasic effect on bone in 2 dif-
this speculation comes from work by Fotsis et al (167) who found
ferent models of ovariectomized rats, young growing rats and lac-
that at high concentrations (IC , 150 mol), genistein inhibited
tating rats, both of which were fed low-calcium diets. These stud-
the ability of bovine microvascular cells to invade collagen gels
ies used 3 different doses of genistein: 1.0, 3.2, and 10 mg/d. After
and generate capillary-like structures when stimulated by basic
2 wk of treatment for the young growing rats and 5 wk of treatment
fibroblast growth factor. Development of antiangiogenesis agents
for the lactating rats, genistein at the lowest dose helped to prevent
is a highly promising area of cancer treatment because inhibiting
ovariectomy-induced, bone-related changes to an extent similar to
the tumor-stimulated growth of new blood vessels prevents
the effects of conjugated equine estrogens (5 g/d).
tumors from becoming larger than 1–2 mm. Tumors limited to
In 1996, Arjmandi et al (98) studied the effects of soy protein
this size are clinically insignificant (168). The concentration of
on bone loss due to ovariectomy. Sprague-Dawley rats were
genistein required to inhibit angiogenesis in vitro, as reported ini-
divided into 4 groups: 1) sham operated, 2) ovariectomized plus
tially (167), is certainly much higher than the genistein concen-
casein, 3) ovariectomized plus soy (0.227 g/g diet, isoflavone
tration likely to be achieved in vivo. However, it has since been
content not indicated), and 4) ovariectomized plus estrogen. The
reported that a much lower genistein concentration is required for
bone density of the right femur was highest in the group given
angiogenesis inhibition in vitro (IC , 8 mol) (94), and that the
estrogen and lowest in the ovariectomized animals fed casein.
initial higher concentration was a result of incomplete solubiliza-
The bone density of the soy group was significantly lower than
that of the estrogen and sham groups, but significantly higher
There is some preliminary support from in vivo research for
than that of the ovariectomized group fed casein. Bone density of
the antiangiogenic potential of genistein. In a small study of
the fourth lumbar vertebra of the soy group was equal to that of
patients with hereditary hemorrhagic telangiectasia, soy intake
the estrogen group and significantly higher than that of both the
led to a marked reduction in nosebleeds and gastrointestinal bleed-
casein and sham groups. This suggests that soy is more protec-
ing (JR Korzenik, S Barnes, unpublished observations, 1996). A
tive of trabecular bone than cortical bone. Similar conclusions
were reached by Anderson et al (180). In a follow-up study by
also worth noting that estrogen and tamoxifen, both of which
Arjmandi et al (181), in which a similar experimental model as
inhibit bone resorption, cause osteoclast apoptosis, an effect that
described above (98) was used, a soy product low in isoflavones
is inhibited in vitro by the addition of antibodies to TGF (188).
did not affect bone density favorably but a soy product high in
As noted previously, genistein was shown to increase TGF in
isoflavones did, clearly suggesting that the isoflavones are
vitro, and thus an effect of genistein on bone resorption may be
responsible for these beneficial effects of soy.
Two other rat studies suggest that genistein in particular affects
The relation between isoflavones and bone health is provoca-
bone density (182, 183). Blair et al (182) fed ovariectomized rats
tive. Thus far, no long-term human studies have examined the
an AIN-76 diet (ICN Pharmaceuticals Inc, Cleveland) or the same
effects of either soy or isoflavones on bone density or even mark-
diet containing 30 mol genistein/d for 4 wk. The dry femoral
ers of bone formation and resorption, let alone fracture risk.
mass of the animals fed genistein was 12% higher (P < 0.05) than
Consequently, although the effect of soy and isoflavones on bone
that of the controls. In a study by Fanti et al (183), after 21 d of
health constitutes an exciting area of research, no firm conclu-
treatment with genistein in ovariectomized rats, 5 and 25 g genis-
sions can be reached at this time. Fortunately, because of the
tein/g body wt injected subcutaneously significantly reduced
number of studies underway, it is likely that a much better under-
ovariectomized tibial bone mineral loss; however, 1 g genistein/g
standing of this issue will be obtained within a relatively short
In contrast to the favorable effects observed in rat studies (98,
180–183), Jayo et al (100) found that in ovariectomized cyno-mogulus monkeys, feeding diets containing soy with or without
SUMMARY AND CONCLUSIONS
isoflavones for 23 mo did not retard the loss of lumbar spine
Legumes have traditionally been an important part of the diets
bone mineral content, whereas monkeys given conjugated equine
of many cultures throughout the world. In contrast, in developed
estrogens had an increase in bone mineral content during this
countries beans currently have only a minor dietary role. The
period. Also, in rats a diet containing an amount of soy that
nutritional profile of beans shows that they have much to offer;
retarded ovariectomy-induced bone loss when administered
beans are high in protein, low in saturated fat, and high in complex
immediately after surgery had no effect when diet administration
carbohydrates and fiber. Beans are also a good source of several
was delayed until 35 d after ovariectomy (184). The implications
micronutrients and phytochemicals. Soybeans are unique among
of this finding may be quite significant given that recent research
the legumes because they are a concentrated source of isoflavones.
suggests that estrogen can exert favorable effects on bone density
It has been hypothesized that isoflavones reduce the risk of cancer,
even when administration is delayed for many years after
heart disease, and osteoporosis, and also help relieve menopausal
symptoms. Although there is much to learn about the effects of
Two human studies that examined the effects of soy consump-
isoflavones on chronic disease risk, this area of research holds
tion on bone mineral loss in postmenopausal women have been
considerable potential. Given the nutrient profile and phytochem-
reported thus far (186, 187). In both studies, soy was associated
ical contribution of legumes, nutritionists should make a concerted
with favorable effects on bone density or content; however, the
effort to encourage the public to consume more beans in general
results of these studies should be considered preliminary. Potter
et al (186) reported that after 6 mo of treatment, lumbar spinebone mineral density increased significantly compared with base-line values in postmenopausal women who consumed 40 g soy
REFERENCES
protein containing 2.25 mg isoflavones/g protein daily, whereas
1. Hellendoorn EW. Beneficial physiologic action of beans. J Am Diet
bone density remained essentially the same in women who con-
sumed the same amount of soy protein but containing only
2. Committee on Diet and Health, National Research Council. Diet
1.39 mg isoflavones/g protein. Women who consumed 40 g of a
and health: implications for reducing chronic disease risk. Washing-
mixture of casein and nonfat dry milk lost bone mineral density
ton, DC: National Academy Press, 1989.
(186). Dalais et al (187) found that early postmenopausal women
3. Life Sciences Research Office, Federation of American Societies for
had a 5% increase in bone mineral content compared with base-
Experimental Biology. Third report on nutrition monitoring in the
line values after only 3 mo of consuming soy flour. Not only
United States. Vol 1. Washington, DC: US Government Printing
does the magnitude of this increase raise questions about these
4. Commercial Agricultural Division, Economic Research Service, US
findings, but the control subjects, who were fed wheat protein,
Department of Agriculture. Dry edible beans: US per capita use for
also experienced an increase in bone mineral content which is
selected classes, 1970–1997. Washington, DC: US Government
surprising given that all the subjects were early postmenopausal
5. US Department of Agriculture. The food guide pyramid. Hyattsville,
Some insight has been gained into the possible mechanism(s)
MD: Human Nutrition Information Service, 1992. (Publication
underlying the effect of isoflavones on bone health in rats. There
are data suggesting that isoflavones may both stimulate and
6. Donovan UM, Gibson RS. Dietary intakes of adolescent females con-
inhibit bone formation. For example, Fanti et al (183) found that
suming vegetarian, semi-vegetarian, and omnivorous diets. J Adolesc
genistein increased osteoblast numbers and serum osteocalcin
concentrations, but had no effect on osteoclast numbers. Con-
7. Ridout CL, Wharf G, Price KR, Johnson LT, Fenwick GR. UK mean
versely, Blair et al (182) studied the effects of genistein on avian
daily intakes of saponins—intestine-permeabilizing factors in
osteoclasts in vitro and found that osteoclast protein synthesis
legumes. Food Sci Nutr 1988;42F:111–6.
was significantly inhibited, an effect that might be due to the
8. Whitten C, Haddad E, Sabaté J. Developing a vegetarian food guide
inhibitory effects of genistein on tyrosine phosphorylation. It is
pyramid: a conceptual framework. Vegetarian Nutr 1997;1:25–9.
9. Sabaté J. Nut consumption, vegetarian diets, ischemic heart disease
32. Simopoulos AP. Essential fatty acids in health and chronic disease.
risk, and all-cause mortality: evidence from epidemiologic studies.
Am J Clin Nutr 1999;70(suppl):560S–9S.
Am J Clin Nutr 1999(suppl);70:500S–3S.
33. Oski FA. What we eat may determine who we can be. Nutrition
10. Kris-Etherton PM, Yu-Poth S, Sabate J, Ratcliffe HE, Zhao G,
Etherton TD. Nuts and their bioactive constituents: effects on serum
34. Emken EA, Adlof RO, Gulley RM. Dietary linoleic acid influences
lipids and other factors that affect disease risk. Am J Clin Nutr
desaturation and acylation of deuterium-labeled linoleic and
linolenic acids in young adult males. Biochim Biophys Acta 1994;
11. Kushi LH, Meyer KA, Jacobs DR Jr. Cereals, legumes, and chronic
disease risk: evidence from epidemiologic studies. Am J Clin Nutr
35. Indu M, Ghafoorunissa. N-3 fatty acids in Indian diets—comparison
of the effects of precursor (alpha-linolenic acid) vs product (long
12. Anderson JW, Smith BS, Washnock CS. Cardiovascular and renal
chain n-3 polyunsaturated fatty acids). Nutr Res 1992;12:569–82.
benefits of dry beans and soybean intake. Am J Clin Nutr
36. Messina M, Messina V. The dietitian’s guide to vegetarian diets: issues
and applications. Rockville, MD: Aspen Publishers, 1996:443–4.
13. Pennington, JAT. Bowes and Churches food values of portions com-
37. WHO and FAO joint consultation: fats and oils in human nutrition.
monly used. 16th ed. Philadelphia: JB Lippincott, 1994.
14. Consumer and Food Economic Institute. Composition of foods: raw,
38. Daly LE, Kirke PN, Molloy A, Weir DG, Scott JM. Folate levels and
processed, and prepared. Agriculture handbook no. 8. Washington,
neural tube defects. JAMA 1995;274:1698–702.
DC: US Government Printing Office, 1976.
39. Lynch SR, Beard JL, Dassenko SA, Cook JD. Iron absorption from
15. National Research Council. Recommended dietary allowances. 10th
legumes in humans. Am J Clin Nutr 1984;40:42–7.
ed. Washington, DC: National Academy Press, 1989.
40. Monsen ER, Hallberg L, Layrisse M, et al. Estimation of available
16. Sarwar G, Peace RW, Botting HG, Brule D. Relationship between
dietary iron. Am J Clin Nutr 1978;31:134–41.
amino acid scores and protein quality indices based on rat growth.
41. Hunt JR, Gallagher SK, Johnson LK. Effect of ascorbic acid on
Plant Foods Hum Nutr 1989;39:33–44.
apparent iron absorption by women with low iron stores. Am J Clin
17. Sarwar G, Peace RW, Botting HG. Corrected relative net protein ratio
(CRNPR) method based on differences in rat and human require-
42. Cook JD, Dassenko SA, Lynch SR. Assessment of the role of non-
ments for sulfur amino acids. J Am Oil Chem Soc 1985;68:689–93.
heme-iron availability in iron balance. Am J Clin Nutr 1991;54:
18. Food and Drug Administration, Department of Health and Human
Services. Food labeling: general requirements for health claims for
43. Sandström B, Almgren A, Kivistö B, Cederblad A. Effect of protein
food. Fed Regist 1991;56:60537–66.
level and protein source on zinc absorption in humans. J Nutr
19. Sarwar G, McDonough FE. Evaluation of protein digestibility-cor-
rected amino acid score method for assessing protein quality of
44. Weaver CM, Heaney RP, Proulz WR, Hinders SM, Packard PT.
foods. J Assoc Off Anal Chem 1990;73:347–56.
Absorbability of calcium from common beans. J Food Sci 1993;58:
20. Sarwar G. The protein digestibility-corrected amino acid score
method overestimates quality of proteins containing antinutritional
45. Weaver CM, Plawecki KL. Dietary calcium: adequacy of a vegetar-
factors and of poorly digestible proteins supplemented with limiting
ian diet. Am J Clin Nutr 1994;59(suppl):1238S–41S.
amino acids in rats. J Nutr 1997;127:758–64.
46. Marlett JA. Content and composition of dietary fiber in 117 fre-
21. Chan JCM. The influence of dietary intake on endogenous acid pro-
quently consumed foods. J Am Diet Assoc 1992;92:175–86.
47. Anderson JW, Story L, Sieling B, Chen W-JL. Hypocholesterolemic
22. Remer T, Manz F. Estimation of the renal net acid excretion by
effects of high-fibre diets rich in water-soluble plant fibres. J Can
adults consuming diets containing variable amounts of protein. Am
48. Jenkins DJA, Wolever TMS, Taylor RH, Barker HM, Fielden H.
23. Kerstetter JE, Allen LH. Dietary protein increases urinary calcium.
Exceptionally low blood glucose response to dried beans: compari-
son with other carbohydrate rich foods. Br Med J 1980;281:578–80.
24. Anderson JJB, Thomsen K, Christiansen, C. High protein meals,
49. Foster-Powell K, Miller JB. International tables of glycemic index.
insular hormones and urinary calcium excretion in human subjects.
Am J Clin Nutr 1995(suppl);62:871S–90S.
In: Christiansen C, Johansen JS, Riis BJ, eds. Osteoporosis. Viborg,
50. Thorne MJ, Thompson LU, Jenkins DJ. Factors affecting starch
Denmark: Nørhaven A/S, 1987:240–5.
digestibility and the glycemic response with special reference to
25. Breslau NA, Brinkley L, Hill KD, Pack CYC. Relationship of ani-
legumes. Am J Clin Nutr 1983;38:481–8.
mal protein-rich diet to kidney stone formation and calcium metab-
51. Thompson LU, Yoon JH, Jenkins DJ, Wolever TM, Jenkins AL.
olism. J Clin Endocrinol Metab 1988;66;140–6.
Relationship between polyphenol intake and blood glucose response
26. Kerstetter JE, O’Brien KO, Insogna KL. Dietary protein influences
of normal and diabetic individuals. Am J Clin Nutr 1984;39:745–51.
intestinal calcium absorption. Am J Clin Nutr 1997;66:215 (abstract 70).
52. Yoon JH, Thompson LU, Jenkins DJ. The effect of phytic acid on in
27. Pannemans DLE, Schaafsma G, Westerterp KR. Calcium excretion,
vitro rate of starch digestibility and blood glucose response. Am J
apparent calcium absorption and calcium balance in young and
elderly subjects: influence of protein intake. Br J Nutr 1997;77:721–9.
53. Nutrition recommendations and principles for people with diabetes
28. US Department of Agriculture, Nutrient Data Research Branch,
mellitus. Diabetes Care 1994;17:519–22.
Nutrition Monitoring Division. Provisional table on the content of
54. Wolever TMS. The glycemic index: flogging a dead horse? Diabetes
omega-3 fatty acids and other fat components in selected foods.
Hyattsville, MD: Human Nutrition Information Service, 1988.
55. Salmerón J, Manson JE, Stampfer MJ, Colditz GA, Wing AL, Wil-
let WC. Dietary fiber, glycemic load, and risk of non-insulin depen-
29. Nair SSD, Leitch JW, Falconer J, Garg ML. Prevention of cardiac
dent diabetes mellitus in women. JAMA 1997;277:472–7.
arrhythmia by dietary (nϪ3) polyunsaturated fatty acids and their
56. Grant G. Anti-nutritional factors of soybean: a review. Prog Food
mechanism of action. J Nutr 1997;127:383–93.
30. Stone NJ. Fish consumption, fish oil, lipids, and coronary heart dis-
57. Duarte-Rayas P, Bergeron D, Nielsen SS. Screening of heat-stable
ease. Am J Clin Nutr 1997;65:1083–6.
trypsin inhibitors in dry beans and their partial purification from great
31. Caygill CPJ, Charlett A, Hill MJ. Fat, fish, fish oil and cancer. Br J
Northern beans (Phaseolus vulgaris) using anhydrotrypsin-sepharose
affinity chromatography. J Agric Food Chem 1992;40:32–42.
58. Liener IE. Implications of antinutritional components in soybean
82. Kelly GE, Joannu GE, Reeder AY, Nelson C, Waring MA. The vari-
foods. Crit Rev Food Sci Nutr 1994;34:31–67.
able metabolic response to dietary isoflavone in humans. Proc Soc
59. Kennedy AR, Manzone H. Effects of protease inhibitors on levels of
proteolytic activity in normal and premalignant cells and tissues.
83. Bradbury RB, White DE. Oestrogens and related substances in
J Cell Biochem 1995;22(suppl):188–94.
plants. Vitam Horm 1954;12:207–12.
60. Oberleas D, Harland BE. Phytate content of foods: effect on dietary
84. Folman Y, Pope GS. The interaction in the immature mouse of potent
zinc bioavailability. J Am Diet Assoc 1981;79:433–6.
oestrogens with coumestrol, genistein and other utero-vaginotrophic
61. Mage JA. Phytate: its chemistry, occurrence, food interactions,
compounds of low potency. J Endocrinol 1966;34:215–25.
nutritional significance, and methods of analysis. J Agric Food
85. Geynet C, Millet C, Truong H, Baulieu EE. Estrogens and antie-
strogens. Hormone antagonists. Gynecol Invest 1972;3:2–29.
62. Graf E, Eaton JW. Antioxidant functions of phytic acid. Free Radic
86. Martin PM, Horwitz KB, Ryan DS, McGuire WL. Phytoestrogen
interaction with estrogen receptors in human breast cancer cells.
63. Harland BF, Morris ER. Phytate: a good or a bad food component?
87. Biggers JD, Curnow DH. Oestrogenic activity of subterranean
64. Vucenik I, Yang G-Y, Shamsuddin AM. Comparison of pure inositol
hexaphosphate and high-bran diet in the prevention of DMBA-
88. Bickoff EM, Livingston AL, Hendrickson AP, Booth AN. Relative
induced rat mammary carcinogenesis. Nutr Cancer 1997;28:7–13.
potencies of several estrogen-like compounds found in forages.
65. Steggerda FR, Dimmick JF. Effects of bean diets on concentration
of carbon dioxide in flatus. Am J Clin Nutr 1966;19:120–4.
89. Mayr U, Butsch A, Schneider S. Validation of two in vitro test sys-
66. Rackis JJ, Sessa DJ, Steggerda FR, Shimizu T, Anderson T, Pearl
tems for estrogenic activities with zearalenone, phytoestrogens and
SL. Soybean factors relating to gas production by intestinal bacte-
cereal extracts. Toxicology 1992;74;135–49.
90. Markiewicz L, Garey J, Adlercreutz H, Gurpide E. In vitro bioas-
67. Carlsson N-G, Karlsson H, Sandberg A-S. Determination of
says of non-steroidal phytoestrogens. J Steroid Biochem Mol Biol
oligosaccharides in foods, diets, and intestinal contents by high-
temperature gas chromatography and gas chromatography/mass
91. Xu X, Harris KS, Wang H-J, Murphy PA, Hendrich S. Bioavailabil-
spectrometry. J Agric Food Chem 1992;40;2404–12.
ity of soybean isoflavones depends upon gut microflora in women.
68. Kuo TM, VanMiddlesworth JF, Wolf WJ. Content of raffinose
oligosaccharides and sucrose in various plant seeds. J Agric Food
92. Adlercreutz H, Markkanen H, Watanabe S. Plasma concentra-
tions of phyto-oestrogens in Japanese men. Lancet 1993;
69. Anderson RL, Rackis JJ, Tallent WH. Biologically active substances
in soy products. In: Wilcke HL, Hopkins DT, Waggle DH, eds. Soy
93. Mäkela SI, Pylkkänen LH, Santti RSS, Adlercreutz H. Dietary soy-
protein and human nutrition. New York: Academic Press, 1979.
bean may be antiestrogenic in male mice. J Nutr 1995;125:437–45.
70. Mitsuoka T. Recent trends in research on intestinal flora. Bifi-
94. Adlercreutz H, Mazur W. Phyto-oestrogens and western diseases.
dobacteria Microflora 1982;1:3–24.
71. Benno Y, Endo K, Mizutani T, Namba Y, Komori T, Mitsuoka T.
95. Santell RC, Chang YC, Nair MG, Helferich WG. Dietary genistein
Comparison of fecal microflora of elderly persons in rural and urban
exerts estrogenic effects upon the uterus, mammary gland and the
areas of Japan. Appl Environ Microbiol 1989;55:1100–5.
hypothalmic/pituitary axis in rats. J Nutr 1997;127:263–9.
72. Koo M, Rao AV. Long term effect of bifidobactaria and neosugar on
96. Dodge JA, Glasebrook AL, Magee DE, et al. Environmental estro-
precursor lesions. Nutr Cancer 1991;16:249–57.
gens: effects on cholesterol lowering and bone in ovariectomized
73. Hata Y, Yamamoto M, Nakajima K. Effects of soybean oligosaccha-
rat. J Steroid Biochem Mol Biol 1996;59:155–61.
rides on human digestive organs: estimate of fifty percent effective
97. Markaverich BM, Webb B, Densmore CL, Gregory RR. Effects of
dose and maximum non-effective dose based on diarrhea. J Clin
coumestrol on estrogen receptor function and uterine growth in
ovariectomized rats. Environ Health Perspect 1995;103:574–81.
74. Slavin JL, Martini MC, Jacobs DR Jr, Marquart L. Plausible mech-
98. Arjmandi BH, Alekel L, Hollis BW, et al. Dietary soybean protein
anisms for protectiveness of whole grains. Am J Clin Nutr
prevents bone loss in an ovariectomized rat model of osteoporosis.
75. Milgate J, Roberts DCK. The nutritional and biological significance
99. Anthony MS, Clarkson TB, Hughes CL Jr, Morgan TM, Burke GL.
of saponins. Nutr Res 1995;15:1223–49.
Soybean isoflavones improve cardiovascular risk factors without
76. Koratkar R, Rao AV. Effect of soya bean saponins on azoxymethane-
affecting the reproductive system of peripubertal rhesus monkeys.
induced preneoplastic lesions in the colon of mice. Nutr Cancer
100. Jayo MJ, Anthony MS, Register TC, Rankin SE, Vest T, Clarkson
77. Coward L, Barnes NC, Setchell KDR, Barnes S. Genistein,
TB. Dietary soy isoflavones and bone loss in ovariectomized mon-
daidzein, and their -glycoside conjugates: antitumor isoflavones in
soybean foods from American and Asian diets. J Agric Food Chem
101. Collins BM, McLachlan JA, Arnold SF. The estrogenic and antie-
strogenic activities of phytochemicals with the human estrogen
78. Wang H-J, Murphy PA. Isoflavone content in commercial soybean
receptor expressed in yeast. Steroids 1997;62:365–72.
foods. J Agric Food Chem 1994;42:1666–73.
102. Loukovaara M, Carson M, Palotie A, Adlercreutz H. Regulation of
79. Wang H-J, Murphy PA. Isoflavone composition of American and
sex hormone-binding globulin production by isoflavonoids and pat-
Japanese soybeans in Iowa: effects of variety, crop year, and loca-
terns of isoflavonoid conjugation in HepG2 cell cultures. Steroids
tion. J Agric Food Chem 1994;42:1674–7.
80. Joannou GE, Kelly GE, Reeder AY, Waring M, Nelson C. A urinary
103. Cassidy A, Bingham S, Setchell KD. Biological effects of a diet of
profile study of dietary phytoestrogens. The identification and mode
soy protein rich in isoflavones on the menstrual cycle of pre-
of metabolism of new isoflavonoids. J Steroid Biochem Mol Biol
menopausal women. Am J Clin Nutr 1994;60:333–40.
104. Cassidy A, Bingham S, Setchell KD. Biological effects of iso-
81. Setchell KD, Borriello SP, Hulme P, Kirk DN, Axelson M. Nons-
flavones in young women: importance of the chemical composition
teroidal estrogens of dietary origin: possible roles in hormone-
of soybean products. Br J Nutr 1995;74:587–601.
dependent disease. Am J Clin Nutr 1984;40:569–78.
105. Baird DD, Umbach DM, Lansdell L, et al. Dietary intervention
study to assess estrogenicity of dietary soy among postmenopausal
127. Rauth S, Kichina J, Green A. Inhibition of growth and induction of
women. J Clin Endocrinol Metab 1995;80:1685–90.
differentiation of metastatic melanoma cells in vitro by genistein:
106. Wang TTY, Sathyamoorthy N, Phang JM. Molecular effects of
chemosensitivity is regulated by cellular p53. Br J Cancer 1997;75:
genistein on estrogen receptor mediated pathways. Carcinogenesis
128. Akiyama T, Ogawara H. Use and specificity of genistein as inhibitor
107. McDonnell DP, Norris JD. Analysis of the molecular pharmacology
of protein-tyrosine kinases. Methods Enzymol 1991;201:362–70.
of estrogen receptor agonists and antagonists provides insights into
129. Constantinou A, Huberman E. Genistein as an inducer of tumor cell
the mechanism of action of estrogen in bone. Osteoporos Int
differentiation: possible mechanisms of action. Proc Soc Exp Biol
108. Kuiper GGJM, Carlsson B, Grandien K, et al. Comparison of the
130. Scholar EM, Toewa ML. Inhibition of invasion of murine mammary
ligand binding specificity and transcript tissue distribution of estro-
carcinoma cells by the tyrosine kinase inhibitor genistein. Cancer
gen receptors ␣ and . Endocrinology 1997;138:863–70.
109. Wrensch MR, Petrakis NL, King EB, et al. Breast cancer incidence
131. Santibáñez JF, Navarro A, Martinez J. Genistein inhibits prolifera-
in women with abnormal cytology in nipple aspirates of breast fluid.
tion and in vitro invasive potential of human prostatic cancer cell
lines. Anticancer Res 1997;17:1199–1204.
110. Petrakis NL, Barnes S, King EB, et al. Stimulatory influence of soy
132. Wei H, Wei L, Frenkel K, Bowen R, Barnes S. Inhibition of tumor
protein isolate on breast fluid secretion in pre- and postmenopausal
promoter-induced hydrogen peroxide formation in vitro and in vivo
women. Cancer Epidemiol Biomarkers Prev 1996;5:785–94.
by genistein. Nutr Cancer 1993;20:1–12.
111. McMichael-Phillips DF, Harding C, Morton M, et al. Effects of soy-
133. Akiyama T, Ishida J, Nakagawa S, et al. Genistein, a specific
protein supplementation on epithelial proliferation in histologically
inhibitor of tyrosine-specific protein kinases. J Biol Chem 1987;
normal human breasts. Am J Clin Nutr 1998;68(suppl):1431S–6S.
112. Messina M, Persky V, Setchell KDR, Barnes S. Soy intake and can-
134. Thorburn J, Thorburn T. The tyrosine kinase inhibitor, genistein,
cer risk: a review of the in vitro and in vivo data. Nutr Cancer 1994;
prevents ␣-adrenergic-induced cardiac muscle cell hypertrophy by
inhibiting activation of the Ras-MAP kinase signaling pathway.
113. Messina MJ, Barnes S. The role of soy products in reducing risk of
Biochem Biophys Res Commun 1994;202:1586–91.
cancer. J Natl Cancer Inst 1991;83:541–6.
135. Linassier C, Pierre M, Le Peco J-B, Pierre J. Mechanism of action
114. Jing Y, Nakaya K, Han R. Differentiation of promyelocytic
in NIH-3T3 cells of genistein, an inhibitor of EGF receptor tyrosine
leukemia cells HL-60 induced by daidzein in vitro and in vivo. Anti-
kinase activity. Biochem Pharmacol 1990;39:187–93.
136. Constantinou A, Kiguchi K, Huberman E. Induction of differentia-
115. Peterson G, Barnes S. Genistein inhibition of the growth of human
tion and DNA strand breakage in human HL-60 and K-562 leukemia
breast cancer cells: independence from estrogen receptors and the
cells by genistein. Cancer Res 1990;50:2618–24.
multi-drug resistance gene. Biochem Biophys Res Commun 1991;
137. Peterson TG, Kim H, Barnes S. Genistein may inhibit the growth of
human mammary epithelial (HME) cells by augmenting transform-
116. Peterson G, Barnes S. Genistein inhibits both estrogen and growth
ing growth factor beta (TGF) signaling. Am J Clin Nutr 1998;
factor-stimulated proliferation of human breast cancer cells. Cell
138. Benson JR, Colletta AA. Transforming growth factor . Prospects for
117. Pagliacci MC, Smacchia M, Migliorati G, Grignana F, Riccardi C,
cancer prevention and treatment. Clin Immunother 1995;4:249–58.
Nicoletti I. Growth-inhibitory effects of the natural phytoestrogen
139. Benson JR, Baum M, Colletta AA. Role of TGF in the anti-estro-
genistein in MCF-7 human breast cancer cells. Eur J Cancer 1994;
gen response/resistance of human breast cancer. J Mammary Gland
118. Peterson G, Coward L, Kirk M, Falany C, Barnes S. The role of metab-
140. Markowitz SD, Roberts AB. Tumor suppressor activity of the TGF-
olism in mammary epithelial growth inhibition by the isoflavones
 pathway in human cancers. Cytokine Growth Factor Rev
genistein and biochanin A. Carcinogenesis 1996;17:1861–9.
119. So FV, Guthrie N, Chambers AF, Moussa M, Carroll KK. Inhibition
141. Cancer facts and figures. Atlanta: American Cancer Society, 1994.
of human breast cell proliferation by flavonoids and citrus juice.
142. Barnes S, Grubbs C, Setchell KDR, Carlson J. Soybeans inhibit
mammary tumors in models of breast cancer. In: Pariza MW,
120. Clark JW, Santos-Moore A, Stevenson LE, Frackelton AR. Effects
Aeschbacher H-U, Felton JS, Sato S, eds. Mutagens and carcino-
of tyrosine kinase inhibitors on the proliferation of human breast
gens in the diet. New York: Wiley Liss, 1990:239–53.
cancer lines and proteins important in the RAS signaling pathway.
143. Lee HP, Gourley L, Duffy SW, Esteve J, Day NE. Dietary effects on
breast-cancer risk in Singapore. Lancet 1991;337:1197–200.
121. Zava DT, Duwe G. Estrogenic and antiproliferative properties of
144. Hirose K, Tajima K, Hamajima N, et al. A large-scale, hospital-
genistein and other flavonoids in human breast cancer cells in vitro.
based case-control study of risk factors of breast cancers according
to menopausal status. Jpn J Cancer Res 1995;86:146–54.
122. Peterson G, Barnes S. Genistein and biochanin A inhibit the growth
145. Yuan J-M, Wang Q-S, Ross RK, Henderson BE, Yu MC. Diet and
of human prostate cancer cells but not epidermal growth factor
breast cancer in Shanghai and Tianjin, China. Br J Cancer 1995;71:
receptor autophosphorylation. Prostate 1993;22:335–45.
123. Naik HR, Lehr JE, Pienta KJ. An in vitro and in vivo study of anti-
146. Wu AH, Ziegler RG, Horn-Ross PL, et al. Tofu and risk of breast
tumor effects of genistein on hormone refractory prostate cancer.
cancer in Asian-Americans. Cancer Epidemiol Biomarkers Prev
124. Kyle E, Neckers L, Takimoto C, Curt G, Bergan R. Genistein-
147. Greenstein J, Kushi L, Zheng W, et al. Risk of breast cancer associ-
induced apoptosis of prostate cancer cells is preceded by a specific
ated with intake of specific foods and food groups. Am J Epidemiol
decrease in focal adhesion kinase activity. Mol Pharmacol 1997;51:
148. Peterson G, Barnes S. Genistein potently inhibits the growth of
125. Kuo S-M, Morehouse HF Jr, Lin C-P. Effect of antiproliferative
human primary breast epithelial cells: correlation with lack of
flavonoids on ascorbic acid accumulation in human colon adenocar-
genistein metabolism. Mol Biol Cell 1994;5:384a (abstr).
cinoma cells. Cancer Lett 1997;116:131–7.
149. Constantinou AL, Mehta RG, Vaughan A. Inhibition of N-methyl-N-
126. Kuo S-M. Antiproliferative potency of structurally distinct dietary
nitrosourea–induced mammary tumors in rats by the soybean
flavonoids on human colon cancer cells. Cancer Lett 1996;110:41–8.
isoflavones. Anticancer Res 1996;16:3293–8.
150. Evans BAJ, Griffiths K, Morton MS. Inhibition of 5”-reductase in
ment with ipriflavone on bone in postmenopausal women with low
genital skin fibroblasts and prostate tissue by dietary lignans and
bone mass. Calcif Tissue Int 1994;54:377–80.
isoflavonoids. J Endocrinol 1995;147:295–302.
171. Brandi ML. Flavonoids: biochemical effects and therapeutic appli-
151. Franke AA, Mordan LJ, Conney RV, et al. Dietary phenolic agents
cations. Bone Miner 1992;19(suppl):S3–64.
inhibit neoplastic transformation and trap toxic NO. Proc Am Assoc
172. Tsuda M, Kitazaki T, Ito T, Fujita T. The effect of ipriflavone (TC-80)
on bone resorption in tissue culture. J Bone Miner Res 1986;1:207–11.
152. Brown NM, Lamartiniere CA. Xenoestrogens alter mammary gland
173. Ross PD, Norimatsu H, Davis JW, et al. A comparison of hip frac-
differentiation and cell proliferation in the rat. Environ Health Per-
ture incidence among native Japanese, Japanese Americans, and
American Caucasians. Am J Epidemiol 1991;133:801–9.
153. Lamartiniere CA, Moore JB, Brown NM, Thompson R, Hardin MJ,
174. Fujita T, Fukase M. Comparison of osteoporosis and calcium intake
Barnes S. Genistein suppresses mammary cancer in rats. Carcino-
between Japan and the United States. Proc Soc Exp Biol Med 1992;
154. Murrill WB, Brown NM, Zhang J-X, et al. Prepubertal genistein
175. Kin K, Lee JH, Kushida K, et al. Bone density and body composi-
exposure suppresses mammary cancer and enhances gland differen-
tion on the Pacific Rim: a comparison between Japan-born and U.S.-
tiation in rats. Carcinogenesis 1996;17:1451–7.
born Japanese-American women. J Bone Miner Res 1993;8:861–9.
155. Yatani R, Kusano I, Shiraishi T, Hayashi T, Stemmerman GN. Latent
176. Ross PD, Fujiwara S, Huang C, et al. Vertebral fracture prevalence
prostatic carcinoma: pathological and epidemiological aspects. Jpn J
in women in Hiroshima compared to Caucasians or Japanese in the
US. Int J Epidemiol 1995;24:1171–7.
156. Pienta KJ, Esper PS. Risk factors for prostate cancer. Ann Intern
177. Russell-Aulet M, Wang J, Thornton JC, Colt EW, Pierson RN Jr.
Bone mineral density and mass in a cross-sectional study of white
157. Ross RK, Bernstein LA, Lobo RA, et al. 5-Alpha-reductase activity
and Asian women. J Bone Miner Res 1993;8:575–82.
and risk of prostate cancer among Japanese and US white and black
178. Nakamura T, Turner CH, Yoshikawa T, et al. Do variations in hip
geometry explain differences between Japanese and white Ameri-
158. Lu L-J, Anderson KE, Nagamani M. Effects of one month soya con-
cans? J Bone Miner Res 1994;9:1071–6.
sumption on circulating steroids in men. Proc Am Assoc Cancer Res
179. Davis JW, Ross PD, Nevitt MC, Wasnich RD. Incidence rates of
falls among Japanese men and women living in Hawaii. J Clin Epi-
159. Pollard M, Luckert PH. Influence of isoflavones in soy protein iso-
lates on development of induced prostate-related cancers in L-W
180. Anderson JJ, Ambrose WW. Orally dosed genistein from soy and
prevention of cancellous bone loss in two ovariectomized rat mod-
160. Zhang JX, Hallmans G, Landström M, et al. Soy and rye diets
els. J Nutr 1995;125(suppl):799S (abstr).
inhibit the development of Dunning R3327 prostatic adenocarci-
181. Arjmandi BH, Birnbaum R, Goyal NV, et al. Bone-sparing effect of soy
noma in rats. Cancer Lett 1997;114:313–4.
protein in ovarian-hormone–deficient rats is related to its isoflavone
161. Schleicher R, Zheng M, Zhang M, Lamartiniere CA. Genistein inhi-
bition of prostate cancer cell growth and metastasis in vivo. Am J
content. Am J Clin Nutr 1998;68(suppl):1364S–8S.
Clin Nutr 1998;68(suppl):1526S(abstr).
182. Blair HC, Jordon SE, Peterson TG, Barnes S. Variable effects of tyro-
162. Dalu A, Haskell J, Lamartiniere CA. Dietary genistein inhibits pro-
sine kinase inhibitors on avian osteoclastic activity and reduction of
tein tyrosine phosphorylation in the dorsolateral prostate of the rat.
bone loss in ovariectomized rats. J Cell Biochem 1996;61:629–37.
Am J Clin Nutr 1998;68(suppl):1524S(abstr).
183. Fanti O, Faugere MC, Gang Z, Schmidt J, Cohen D, Malluche HH.
163. Geller J, Sionit L, Partido C, et al. Genistein inhibits the growth of
Systemic administration of genistein partially prevents bone loss in
human-patient BPH and prostate cancer in histoculture. Prostate
ovariectomized rats in a nonestrogen-like mechanism. Am J Clin
164. Severson KJ, Nomura AMY, Grove JS, Stemmermann GN. A prospec-
184. Arjmandi BH, Getlinger MJ, Goyal NV, et al. Role of soy protein
tive study of demographics, diet, and prostate cancer among men of
with normal or reduced isoflavone content in reversing bone loss
Japanese ancestry in Hawaii. Cancer Res 1989;49:1857–60.
induced by ovarian hormone deficiency in rats. Am J Clin Nutr
165. Morton MS, Matos-Ferreira A, Abranches-Monteiro L, et al. Meas-
urement and metabolism of isoflavonoids and lignans in human
185. Schneider DL, Barrett-Connor EL, Morton DJ. Timing of post-
male. Cancer Lett 1997;114:145–51.
menopausal estrogen for optimal bone mineral density. JAMA
166. Stephens FO. Phytoestrogens and prostate cancer: possible preven-
tive role. Med J Aust 1997;167:138–40.
186. Potter SM, Baum JA, Teng H, Stillman RJ, Shay NF, Erdman JW Jr.
167. Fotsis T, Pepper M, Adlercreutz H, et al. Genistein, a dietary-derived
Soy protein and isoflavones: their effects on blood lipids and bone
inhibitor of in vitro angiogenesis. Proc Natl Acad Sci U S A 1993;
density in postmenopausal women. Am J Clin Nutr 1998;68(suppl):
168. Folkman J, Klagsbrun M. Angiogenic factors. Science 1987;235;
187. Dalais FS, Rice GE, Bell RJ, et al. Dietary soy supplementation
increases vaginal cytology maturation index and bone mineral con-
169. McAllister KA, Baldwin MA, Thukkani AK, et al. Six novel muta-
tent in postmenopausal women. Am J Clin Nutr 1998;68(suppl):
tions in the endoglin gene in hereditary hemorrhagic telangiectasia
type 1 suggest a dominant-negative effect of receptor function. Hum
188. Hughes DE, Dai A, Tiffee JC, Li HH, Mundy GR, Boyce BF. Estro-
gen promotes apoptosis of murine osteoclasts mediated by TGF-.
170. Valente M, Bufalino L, Castiglione GN, et al. Effects of 1-year treat-
2014 1L LCLD Scholars Program In connection with the Leadership Council on Legal Diversity (LCLD) Pipeline Committee’s 1L Scholar Program, we are pleased to announce the McGuireWoods/Exelon Energy Internship. This partnership between McGuireWoods and Exelon will provide a unique opportunity for the selected 1L law student to work side by side with McGuireWoods and Exelon Legal Departm
Paul Baines Marketing Walk-Through of the Online Resource Centre Part 1: Marketing Fundamentals 1 Marketing Principles and Society 3 Marketing Psychology and Consumer Buying Behaviour 4 Marketing Research and Marketing Information Systems Part 2: Principles of Marketing Management 5 Marketing Strategy 7 Market Development and International Marketing Part 3: The Marke
 Legumes and soybeans: overview of their nutritional profiles andhealth effects1,2
ABSTRACT
Legumes and soybeans: overview of their nutritional profiles andhealth effects1,2
ABSTRACT Beans have long been recognized for their protein content and
value for protein quality. The PDCAASs of most beans are rea-
more recently have been noted for their soluble-fiber content, but
sonably good, although their overall value is reduced somewhat
in general there has been relatively little research and discussion
by their lower digestibility (19). Some types of soy protein prod-
about the nutritional attributes of legumes. The glaring exception
ucts have PDCAASs of close to one, the highest value possible.
Beans have long been recognized for their protein content and
value for protein quality. The PDCAASs of most beans are rea-
more recently have been noted for their soluble-fiber content, but
sonably good, although their overall value is reduced somewhat
in general there has been relatively little research and discussion
by their lower digestibility (19). Some types of soy protein prod-
about the nutritional attributes of legumes. The glaring exception
ucts have PDCAASs of close to one, the highest value possible. also contain the nϪ3 fatty acid, ␣-linolenic acid (28). However,
index were Ϸ40% more likely to develop diabetes than those
because the overall fat content of most beans is so low, the dietary
consuming low-glycemic-index diets, even after controlling for
contribution of beans to ␣-linolenic acid intake is generally minor.
also contain the nϪ3 fatty acid, ␣-linolenic acid (28). However,
index were Ϸ40% more likely to develop diabetes than those
because the overall fat content of most beans is so low, the dietary
consuming low-glycemic-index diets, even after controlling for
contribution of beans to ␣-linolenic acid intake is generally minor.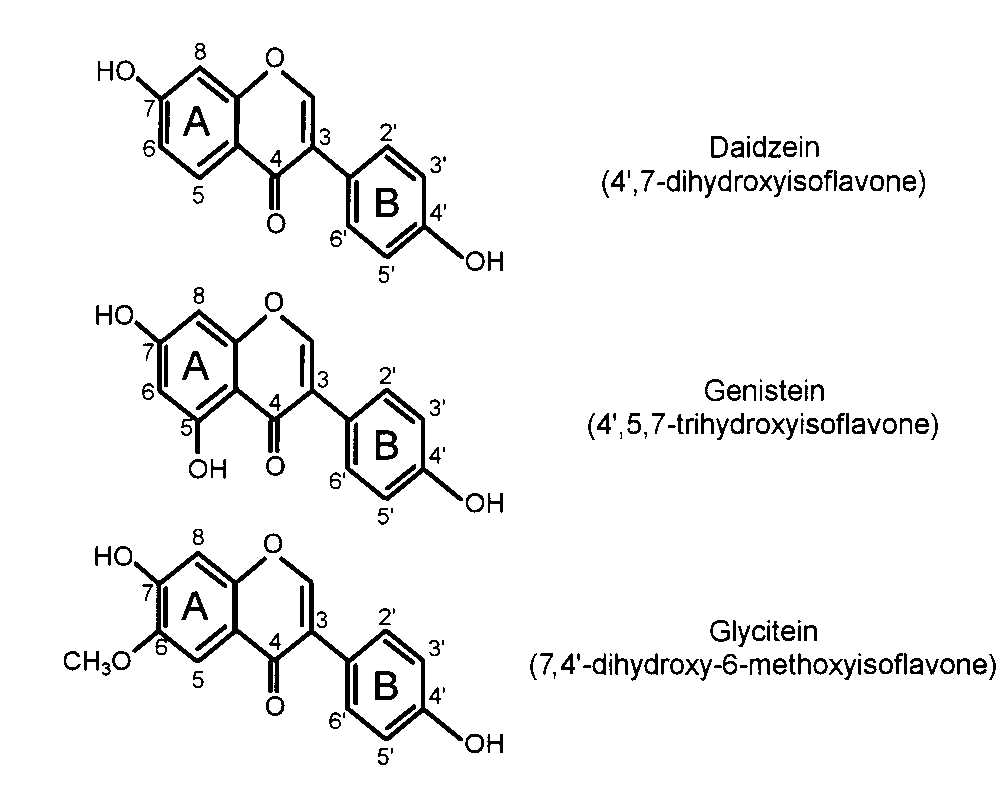
 structure attached to water-soluble sugar residues that differ in
as the conjugate, whereas in fermented soy products such as
their type and amount. The major sources of dietary saponins are
legumes, and many types of saponins can be present in the same
In addition to the isoflavones found in soybeans, the intestinal
bean. Saponins are very poorly absorbed. Most saponins form
microflora can convert daidzein into several different products,
insoluble complexes with 3--hydroxysteroids and are known to
including the isoflavonoids equol (7-hydroxyisoflavan), dihydro-
interact with and form large, mixed micelles with bile acids and
daidzein, and O-desmethylangolensin (80). However, because of
cholesterol. Although saponins were shown to lower cholesterol in
differences in intestinal microflora, equol production occurs in
some animal species, the hypocholesterolemic effects of saponins
only Ϸ1 out of every 3 individuals consuming soyfoods (81, 82).
structure attached to water-soluble sugar residues that differ in
as the conjugate, whereas in fermented soy products such as
their type and amount. The major sources of dietary saponins are
legumes, and many types of saponins can be present in the same
In addition to the isoflavones found in soybeans, the intestinal
bean. Saponins are very poorly absorbed. Most saponins form
microflora can convert daidzein into several different products,
insoluble complexes with 3--hydroxysteroids and are known to
including the isoflavonoids equol (7-hydroxyisoflavan), dihydro-
interact with and form large, mixed micelles with bile acids and
daidzein, and O-desmethylangolensin (80). However, because of
cholesterol. Although saponins were shown to lower cholesterol in
differences in intestinal microflora, equol production occurs in
some animal species, the hypocholesterolemic effects of saponins
only Ϸ1 out of every 3 individuals consuming soyfoods (81, 82). both premenopausal and postmenopausal women taking hormone
addition to the low breast cancer mortality rates in Asia, 2 other
replacement therapy increased in response to soy consumption, as
early observations provided a basis for the hypothesis that soy
did the number of atypical cells in the breast fluid (110). However,
intake decreases breast cancer risk: 1) the potential antiestrogenic
this was a pilot study that did not include a control group.
both premenopausal and postmenopausal women taking hormone
addition to the low breast cancer mortality rates in Asia, 2 other
replacement therapy increased in response to soy consumption, as
early observations provided a basis for the hypothesis that soy
did the number of atypical cells in the breast fluid (110). However,
intake decreases breast cancer risk: 1) the potential antiestrogenic
this was a pilot study that did not include a control group. trations required to inhibit breast cancer–cell growth. It should
soyfoods may be a factor contributing to the low prostate cancer
be noted, however, that Peterson and Barnes (148) found that
mortality rate in Japan, although the data in support of this
genistein inhibits the serum and epidermal growth factor–stimu-
hypothesis, while intriguing, are limited.
trations required to inhibit breast cancer–cell growth. It should
soyfoods may be a factor contributing to the low prostate cancer
be noted, however, that Peterson and Barnes (148) found that
mortality rate in Japan, although the data in support of this
genistein inhibits the serum and epidermal growth factor–stimu-
hypothesis, while intriguing, are limited. way. Related to this finding are those of Geller et al (163), who
larger, follow-up study is currently underway. Interestingly, the
found that genistein (at concentrations of 1–15 g/mL) inhibited
genes in which hereditary hemorrhagic telangiectasia mutations
the incorporation of 3H-thymidine (a measure of tissue growth)
have been mapped thus far all encode for proteins that are involved
in cultured benign prostatic hypertrophy tissue by 44–86% in a
in TGF signaling (169) and as noted previously, Peterson et al
showed that in vitro, genistein increases TGF levels (137).
way. Related to this finding are those of Geller et al (163), who
larger, follow-up study is currently underway. Interestingly, the
found that genistein (at concentrations of 1–15 g/mL) inhibited
genes in which hereditary hemorrhagic telangiectasia mutations
the incorporation of 3H-thymidine (a measure of tissue growth)
have been mapped thus far all encode for proteins that are involved
in cultured benign prostatic hypertrophy tissue by 44–86% in a
in TGF signaling (169) and as noted previously, Peterson et al
showed that in vitro, genistein increases TGF levels (137).
 were reached by Anderson et al (180). In a follow-up study by
also worth noting that estrogen and tamoxifen, both of which
Arjmandi et al (181), in which a similar experimental model as
inhibit bone resorption, cause osteoclast apoptosis, an effect that
described above (98) was used, a soy product low in isoflavones
is inhibited in vitro by the addition of antibodies to TGF (188).
were reached by Anderson et al (180). In a follow-up study by
also worth noting that estrogen and tamoxifen, both of which
Arjmandi et al (181), in which a similar experimental model as
inhibit bone resorption, cause osteoclast apoptosis, an effect that
described above (98) was used, a soy product low in isoflavones
is inhibited in vitro by the addition of antibodies to TGF (188). 9. Sabaté J. Nut consumption, vegetarian diets, ischemic heart disease
32. Simopoulos AP. Essential fatty acids in health and chronic disease.
9. Sabaté J. Nut consumption, vegetarian diets, ischemic heart disease
32. Simopoulos AP. Essential fatty acids in health and chronic disease. 58. Liener IE. Implications of antinutritional components in soybean
82. Kelly GE, Joannu GE, Reeder AY, Nelson C, Waring MA. The vari-
foods. Crit Rev Food Sci Nutr 1994;34:31–67.
58. Liener IE. Implications of antinutritional components in soybean
82. Kelly GE, Joannu GE, Reeder AY, Nelson C, Waring MA. The vari-
foods. Crit Rev Food Sci Nutr 1994;34:31–67. study to assess estrogenicity of dietary soy among postmenopausal
127. Rauth S, Kichina J, Green A. Inhibition of growth and induction of
women. J Clin Endocrinol Metab 1995;80:1685–90.
study to assess estrogenicity of dietary soy among postmenopausal
127. Rauth S, Kichina J, Green A. Inhibition of growth and induction of
women. J Clin Endocrinol Metab 1995;80:1685–90. 150. Evans BAJ, Griffiths K, Morton MS. Inhibition of 5”-reductase in
ment with ipriflavone on bone in postmenopausal women with low
genital skin fibroblasts and prostate tissue by dietary lignans and
bone mass. Calcif Tissue Int 1994;54:377–80.
150. Evans BAJ, Griffiths K, Morton MS. Inhibition of 5”-reductase in
ment with ipriflavone on bone in postmenopausal women with low
genital skin fibroblasts and prostate tissue by dietary lignans and
bone mass. Calcif Tissue Int 1994;54:377–80.