Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Hibernation 28-37 mammals.fh1
Seasonal timing and pre-hibernation
32 fattening in breeding and non-breeding European Ground Squirrels
EVA MILLESI, ANITA DIVJAK & ANNA STRAUSSDepartment of Behavioural Biology, University of Vienna, Althanstr. 14, 1090 Vienna, Austria; eva.millesi@univie.ac.at
Abstract We compared seasonal timing, body mass changes and plasma progesterone secretion in
breeding and non-breeding female European Ground Squirrels. One group of females could
mate normally in spring, other individuals had no access to males. The ground squirrels
were born and kept in semi-natural conditions in outdoor enclosures. Females were captured
at weekly intervals, weighed and reproductive status was determined. The results showed
that the course and extent of prehibernation fattening were similar in non-reproductive
and postreproductive females. In non-breeding females, however, fattening occurred earlier,
resulting in immergence into the hibernacula 46 weeks ahead of the annual schedule. Body
mass at immergence was similar in both groups. Progesterone secretion increased in both
groups prior to pre-hibernation fattening. Before hibernation onset, progesterone secretion
decreased to baseline levels in al females. In general, our results demonstrate that the lack
of reproductive effort in female ground squirrels led to a shift in the seasonal timing of
fattening, earlier immergence and probably an extended hibernation period. Introduction Similar to many other hibernating ground squirrel species, female European Ground Squirrels
(Spermophilus citel us) mate shortly after spring emergence (reviewed in Michener 1984; Mil esi
et al. 1999a), give birth after 4 weeks of gestation and lactate their offspring for 46 weeks. After
weaning, females use the remaining part of their active season to prepare for hibernation.
European Ground Squirrels rely solely on their body fat reserves during winter (Millesi et al.
1998). Thus, the postreproductive period is characterised by a rapid body mass increase during
prehibernation fattening (Mil esi et al. 1999b). Adult females usual y start hibernation in early
August, fol owed by adult males in late August, and juveniles in late September. Female S. citel us
start breeding after their first hibernation and can live for up to six years in the field (Hoffmann
et al. 2003). Hence, a non-reproductive season may on the one hand reduce lifetime reproductive
success, but on the other hand, could enhance future reproductive output by improving a females
Lovegrove, B.G. & McKechnie, A.E. (eds) Hypometabolism in animals: hibernation, torpor and cryobiology. University of KwaZulu-Natal, Pietermaritzburg.
condition in the subsequent spring, as has been shown in alpine marmots (Hackländer & Arnold
1999). In this study we compared the duration of the active season, timing and course of pre-
hibernation fattening, immergence into the hibernaculum, body mass changes and progesterone
secretion in breeding and non-breeding female European Ground Squirrels kept in semi-natural
conditions. Considering the energetic costs of reproductive effort, we expected the non-breeding
females to increase body mass throughout the season, entering hibernation with a higher body
mass than reproductive individuals. Progesterone secretion has been found to increase during
late lactation and remain elevated until shortly before hibernation, indicating an active luteal
phase during the postreproductive period (Strauss et al. unpubl. data). In fact, active corporalutea have been found in females shortly after weaning and during prehibernation fattening
(Mil esi et al. unpubl. data). In this study we compared the course of plasma progesterone secretion
between breeding and non-breeding females. Owing to the absence of pregnancy and lactation
we expected lower progesterone levels in non-pregnant compared to pregnant females and,
similar to the postreproductive progesterone peak mentioned above, elevated progesterone
secretion in non-breeding females during an active luteal phase following ovulation. Materials and methods Animals, study area and capture techniques Female European Ground Squirrels (Spermophilus citel us), living under semi-natural conditions
in outdoor enclosures built within the habitat of the species north of Vienna (Austria), were
investigated from vernal emergence until the onset of hibernation. In the largest enclosure
(240 m2) al males were removed prior to female emergence. Hence, none of these females mated
or showed signs of gestation or lactation (n = 10). In two other enclosures (160 m2 each), females
had access to males and reproduced successfully (n = 8). Individual density was similar in all
enclosures. The density in the enclosures was higher than that of free-living populations in close
distance but corresponded to that in preferred parts during high-density periods (Hoffmann
et al. 2003). Al animals were permanently marked with subcutaneous transponder chips injected
in the neck region (PIT-tag, Datamars Comp.). For recognition from a distance, the fur was
painted with commercial hair dye in a unique pattern. Females were captured at weekly intervals
using Tomahawk live traps baited with peanut butter. At each capture, animals were weighed
(± 0.5 g, Sartorius laboratory scale) and reproductive status was determined. To determine
vernal emergence dates of individual females, all burrows in the enclosures were controlled at
daily intervals between mid-February until al ground squirrels had started aboveground activity.
Opened burrows could easily be detected by digging traces. The animals could be identified by
their fur marks and were captured within one or two days. Gestation and lactation were
determined by examining body-mass changes and teat development. Pregnant females were
recognised by dark-pigmented teats and rapid body mass increase. Parturition was detected
by an abrupt drop of body mass at the expected time. During lactation, females had large,
light-pigmented teats, and milk secretion could be induced. Weaning had occurred when the
teats were dried up and no further signs of lactation were determined. Pre-hibernation fattening
was defined by mass increases < 1 g/d, excluding the gestation period in breeding females (Mil esi
Seasonal timing and pre-hibernation fattening in breeding and non-breeding European Ground Squirrels
Immergence date was determined by the last sighting of an individual female during
summer. In addition, the burrows of the experimental females were frequently checked and
closed burrows of animals that had terminated above-ground activity could be identified.
All animal procedures were performed in accordance with Austrian legal requirements
for animal experimentation and were approved by the Austrian Committee for Animal Research
of the Federal Ministry of Agriculture, Forestry, Environment and Water Management
Hormone assay Blood samples (150200 l) were taken from the femoral vein at weekly intervals. The blood was
collected in heparinised capillaries and centrifuged in the field. Plasma was stored at -20°C
until assayed for hormone concentrations. Progesterone levels were analysed in duplicate after
diethylether extraction with a biotin-streptavidin enzyme immunoassay (Palme & Möstl 1994).
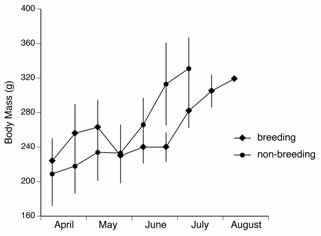
Intra- and inter-assay coefficients of variation were 10.6% and 15.1%. Results Body mass in females that had mated increased during pregnancy in April (Figure 1). At
parturition, body mass dropped and did not increase until weaning in late June. Thereafter,
prehibernation fattening occurred, defined as a period of rapid mass increase (Figure 1). Fattening
started in late June and the body mass increase levelled off by mid-July. In non-breeding
individuals, body mass started to increase rapidly in mid-May. In these individuals fattening
started four weeks earlier than in breeding females (Figure 1). The duration of the fattening
period did not differ significantly between breeding and non-breeding females (breeding: 30.3
± 9.9 d, n = 8; non-breeding: 29 ± 7.1 d, n = 10; Mann-Whitney U test, p = 0.8). Hence, non-
breeding individuals reached their maximum body mass earlier in the season than breeding
females. Daily mass increase rates during fattening were similar in both female groups (breeding:
3.5 ± 1.0 g/d, n = 8; non-breeding: 3.7 ± 1.2 g/d, n = 10; Mann-Whitney U test, p = 0.7).
The differences in the timing of the fattening period were reflected in the termination of
above-ground activity. Non-breeding females immerged about one month earlier than
individuals that had raised a litter (Table 1). Some of the non-breeding females even went into
Figure 1. Body mass changes in
reproductive and non-reproductive EuropeanGround Squirrels. Means
in 2-week intervals ± SDare shown (reproductive:n = 8,8,8,8,8,8,8,8,6,1;non-reproductive:n = 10,10,10,10,10,4,2).
their hibernacula by mid-June. As spring emergence dates did not differ between the two
groups, the active period in non-breeding females was about 4 weeks shorter than in breeding
ones (84 d ± 8 vs 115 d ± 17, p < 0.001, n = 10/8). Neither body mass at vernal emergence nor
shortly before immergence differed significantly between the two female groups (Table 1).
Progesterone secretion patterns differed between the two groups in that during April, levels
were baseline in non-reproductive individuals, but elevated in females that had mated and
become pregnant (Mann-Whitney U test, p < 0.01) (Figure 2). Thereafter, plasma progesterone
secretion increased rapidly in non-breeding females until levels peaked in late May and remained
elevated until mid-June. In breeding females, progesterone levels decreased during parturition
and early lactation, increased thereafter and were elevated from late May until late June. Pro-
gesterone secretion declined earlier in non-breeding females but with the same rate as in breeding
individuals and reached baseline levels in both groups prior to immergence (Figure 2).Discussion Unmated European Ground Squirrels could use additional time for growth and fattening. All
experimental females were older than two years and hence definitely adult. Thus, their
pronounced mass increase was entirely due to fattening. Prehibernatory fattening started earlier
in non-breeding females than in individuals that had raised a litter. Maximum annual body
mass was reached by early July in non-breeding females, about four weeks prior to breeding
individuals. Daily mass increase rates during fattening did not differ in non-reproductive and
postreproductive females.Accordingly, immergence body mass was similar in both groups.
Nunes et al. (2002) showed that juvenile Beldings Ground Squirrels provisioned with food were
Table 1. Dates of emergence and immergence, body mass (g) at emergence and shortly before
immergence in breeding and non-breeding female European Ground Squirrels (means
± SD; n = 8/10; n.s. = not significant). Emergence Date Immergence Date Emergence Date Immergence Date Figure 2. Plasma progesterone secretion patterns in reproductive and non- reproductive European
Ground Squirrels. Meansin 2-week intervals ± SDare shown (reproductive:n = 7,8,8,8,7,8,8,8,6,1;
Seasonal timing and pre-hibernation fattening in breeding and non-breeding European Ground Squirrels
fatter than unprovisioned ones, but that these differences disappeared when prehibernation
fattening started. Accordingly, our results indicate that prehibernation fattening in this species
is a temporally limited process and merely shifted to an earlier period in non-reproductive
females. There may be a certain threshold in body fat content that has to be reached to enable
hibernation, resulting in an earlier and potential y longer hibernation period in non-breeding
females. Early immergence may be beneficial because mortality risk due to predation could
exceed that of an extended hibernation period (Michener 1984). Results of previous field studies
demonstrated that the frequency of observed predators increased rapidly during the first two
weeks of June when the juveniles appeared above ground (Hoffmann et al. unpubl. data).
Although the victims were mainly juveniles, we also observed several kil s of adult individuals.
After weaning, juveniles started to disperse, but the frequency of observed predators only slowly
decreased. This could indicate an increased predation risk for adult individuals in the last part
of the active season. Apparently, unmated ground squirrels did not use the additional time to
get fatter than the others, but preferred to terminate above-ground activity in early summer.
Al females in our enclosures were provided with food, resulting in a good body condition
and sufficient fat reserves. Stil , body mass increase rates of al experimental females were within
the range of free-living European Ground Squirrels (Millesi et al. 1999b). In non-breeding
females, body mass increased very moderately during the first four weeks postemergence. This
could be due to a prolonged oestrous period associated with elevated oestrogen secretion (Mil esi
et al. 2000). Earlier studies had shown that vaginal oestrus in unmated females, defined by the
predominance of cornified cells in vaginal smear samples, lasted for up to two weeks. Oestrus
was terminated by late April, corresponding to the end of the mating period in females with
access to males. The flexibility to prolong oestrus could be adaptive in cases of low population
density or harsh weather conditions in spring (Morton & Sherman 1978; Murie & Harris 1982).
This phenomenon has been found in related species like S. beldingi (Holmes & Landau 1986)
and S. tridecemlineatus (Landau & Holmes 1988) as wel as in Woodchucks (Hikim et al. 1991).
As expected, plasma progesterone levels were significantly higher in pregnant females than
in non-breeding females during the gestation period in April. Similar results were found in
reproductive and non-reproductive Alpine Marmots (Hackländer et al. 2003). Thereafter
progesterone secretion increased rapidly in non-breeding females and remained elevated for
about four weeks. This could be due to an extended active luteal phase following spontaneous
ovulation. Earlier studies have demonstrated a second oestrus cycle after weaning, including
spontaneous ovulation and the formation of an active corpus luteum (Mil esi et al. unpubl. data).
Progesterone secretion increased during late lactation and peaked after weaning in June. The
extended period of progesterone secretion in non-breeding and postbreeding female European
Ground Squirrels seems to be unique among hibernating smal mammals. It may be related to
another unusual phenomenon found in this species: females start hibernation about three weeks
before males, despite the higher energetic and temporal constraints caused by maternal effort
(Mil esi et al. 1999a,b). Elevated progesterone secretion may enhance the progress of prehiberna-
tory fattening. Female rats implanted with progesterone, for example, had an increased pro-
portion of total body fat compared to control animals (Mendes et al. 1984; Shirling et al. 1981).
Moreover, progesterone could increase food intake rates, as has been shown in rats (Shirling
et al. 1981). These effects would be advantageous in animals that have to store body fat in a
limited timespan. Future experimental work is needed to investigate these potential effects.
This study clearly demonstrates that the lack of reproductive effort in female ground squirrels
led to a shift in seasonal timing, an earlier immergence and probably an extended hibernation
Acknowledgements The study was supported by the Austrian Science Fund (FWF, P13646). We thank Anna Schöbitz
for help in carrying out the endocrine analyses. References HACKLÄNDER, K. & ARNOLD, W. 1999. Male-caused failure of female reproduction and its adaptive value in
Alpine marmots (Marmota marmota). Behavioral Ecology and Sociobiology 10:592597.
HACKLÄNDER, K., MÖSTL, E. & ARNOLD, W. 2003. Reproductive suppression in female Alpine marmots,
Marmota marmota. Animal Behaviour 65:11331140.
HIKIM, A.P.S., WOOLF, A., BARTKE, A. & AMADOR, A.G. 1991. The estrous cycle of captive woodchucks (Marmotamonax). Biology of Reproduction 44:733738.
HOFFMANN, I.E., MILLESI, E., EVERTS, L.G., HUBER, S. & DITTAMI, J.P. 2003. Population dynamics of European
ground squirrels (Spermophilus citel us) in a suburban area. Journal of Mammalogy 84:615626.
HOLMES, W.G. & LANDAU, I.T. 1986. Vaginal estrus in unmated Beldings ground squirrels. Hormones and
KROTKIEWSKI, M. & BJÖRNTORP, P. 1976. Effect of progesterone and of insulin administration on regional
adipose tissue cellularity in rats. Acta Physiologica Scandinavica 96:122127.
LANDAU, I.T. & HOLMES, W.G. 1988. Mating of captive thirteen-lined ground squirrels and the annual timing
of estrus. Hormones and Behavior 22:474478.
MENDES, A-M., MADON, R.J. & FLINT, D.J. 1985. Effects of cortisol and progesterone on insulin binding and
lipogenesis in adipocytes from normal and diabetic rats. Journal of Endocrinology 106:225231.
MICHENER, G.R. 1984. Age, sex and species differences in the annual cycle of ground-dwel ing sciurids: implications
for sociality. Pp. 81107 in Murie, J.O. & Michener, G.A. (eds), The biology of ground dwelling squirrels.
University of Nebraska Press, Lincoln.
MILLESI, E., HUBER, S., DITTAMI, J.P., HOFFMANN, I.E. & DAAN, S. 1998. Parameters of mating effort and
success in male European ground squirrels, Spermophilus citel us.Ethology 104:298313.
MILLESI, E., HUBER, S., EVERTS, L.G. & DITTAMI, J.P. 1999a. Reproductive decisions in female European ground
squirrels: factors affecting reproductive output and maternal investment. Ethology 105:163175.
MILLESI, E., STRIJKSTRA, A.M., HOFFMANN, I.E., DITTAMI, J.P. & DAAN, S. 1999b. Sex and age differences
in mass, morphology, and annual cycle in European ground squirrels, Spermophilus citel us.Journal of
MILLESI, E., HUBER, S., PIETA, K., WALZL, M., ARNOLD, W. & DITTAMI, J.P. 2000. Estrus and estrogen changes
in mated and male-deprived free-living European ground squirrels. Hormones and Behavior 37:190197.
MORTON, M.L. & SHERMAN, P.W. 1978. Effects of a spring snowstorm on behavior, reproduction and survival
of Beldings ground squirrels. Canadian Journal of Zoology 56:25782590.
MURIE, J.O. & HARRIS, M.A. 1982. Annual variation of spring emergence and breeding in Columbian ground
squirrels (Spermophilus columbianus). Journal of Mammalogy 63:431439.
NUNES, S., MUECKE, E-M. & HOLECAMP, K.E. 2002. Seasonal effects of food provisioning on body fat, insulin,
and corticosterone in free-living juvenile Beldings ground squirrels (Spermophilus beldingi). CanadianJournal of Zoology 80:366-371.
PALME, R. & MÖSTL, E. 1994. Biotin-streptavidin enzyme immunoassay for the determination of oestrogens and
androgens in boar faeces. Pp. 111-117 in Görög, S. (ed.), Advances in steroid analysis 93. Akadémiai Kiadó,
SHIRLING, D., ASHBY, J.P. & BAIRD, J.D. 1981. Effect of progesterone on lipid metabolism in the intact rat. Journalof Endocrinology 90:285294.
Curriculum Professionale ------------------------------------------------------------------------------------------------------------------------ Luogo e data di nascita: Roma, 06/07/1975 ------------------------------------------------------------------------------------------------------------------------ Luglio 2002 . Laurea in Medicina e Chirurgia presso L’Università degli Studi d
Cimzia® (certolizumab pegol) COVERAGE CRITERIA DRUG CLASS: GI Drugs, Miscellaneous – AHFS 56:92 BRAND NAME: Cimzia® GENERIC NAME: certolizumab pegol POLICY #: 0261 CATEGORY: Commercial FDA INDICATIONS : • Crohn’s Disease (CD) – for reducing signs and symptoms of Crohn’s disease and maintaining clinical response in adult patients with moderately to severely a
 Seasonal timing and pre-hibernation fattening in breeding and non-breeding European Ground Squirrels
Immergence date was determined by the last sighting of an individual female during
summer. In addition, the burrows of the experimental females were frequently checked and
closed burrows of animals that had terminated above-ground activity could be identified.
Seasonal timing and pre-hibernation fattening in breeding and non-breeding European Ground Squirrels
Immergence date was determined by the last sighting of an individual female during
summer. In addition, the burrows of the experimental females were frequently checked and
closed burrows of animals that had terminated above-ground activity could be identified.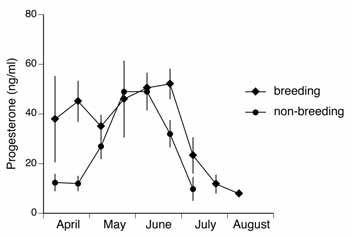 their hibernacula by mid-June. As spring emergence dates did not differ between the two
groups, the active period in non-breeding females was about 4 weeks shorter than in breeding
ones (84 d ± 8 vs 115 d ± 17, p < 0.001, n = 10/8). Neither body mass at vernal emergence nor
shortly before immergence differed significantly between the two female groups (Table 1).
their hibernacula by mid-June. As spring emergence dates did not differ between the two
groups, the active period in non-breeding females was about 4 weeks shorter than in breeding
ones (84 d ± 8 vs 115 d ± 17, p < 0.001, n = 10/8). Neither body mass at vernal emergence nor
shortly before immergence differed significantly between the two female groups (Table 1).