Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Carlosibanezlab.se
Inducible and tissue-specific gene targeting to cholinergic neurons
Division of Molecular Neurobiology, Department of Neuroscience,Karolinska Institute, S-171 77 Stockholm, Sweden
Keywords: gene regulation, transgenic mice, choline acetyltransferase, tetracycline, enhancer,
We have devised a strategy for cell type-specific and regulatable targetedexpression of genes to cholinergic neurons. A 2336 base pair-long fragment from therat choline acetyltransferase (ChAT) gene regulatory region, which we previouslyshowed to direct expression to cholinergic neurons in transgenic mice, was cloned infront of the cytomegalovirus (CMV) and herpes simplex virus thymidine kinase (TK)promoters to control the expression of the rtTA chimeric transcriptional transactivator. Addition of the tetracycline analogue doxycycline transforms rtTA in a potenttransactivator protein, capable of triggering high level of expression from reporterconstructs containing a minimal promoter and binding sites from the tetracyclineoperon. Inducible expression of the luciferase reporter gene was seen in thecholinergic SN6 cell line upon doxycycline treatment. No induction above backgroundwas seen in FR3T3 fibroblasts transfected with the same constructs. The induction b ydoxycycline of reporter gene expression in SN6 cells was dose-dependent andsaturable. Transgenic mice were generated with the ChAT-TK-rtTA construct. One ofthese lines showed predominant expression of rtTA mRNA in the spinal cord, one ofthe richest sources in cholinergic cell bodies in the nervous system. These animals willbe of great value for developmental and functional studies on cholinergic neurons.
Inducible and tissue-specific gene targeting to cholinergic neurons
An important population of neurons in the nervous system are the cholinergic neurons,mainly located in the ventral horn of the spinal cord, cranial nuclei, and in the basalforebrain. Cholinergic cell bodies are also found in cortex, striatum and the olfactorybulb. The cholinergic systems are involved in motor and cognitive functions and areaffected in Alzheimer’s disease. Choline acetyltransferase (ChAT) is the enzymeresponsible for the synthesis of acetylcholine, and, together with the vesicularacetylcholine transporter (VAChT), is the best available marker for cholinergic neurons(Arvidsson et al., 1997). The VAChT gene has been shown to be contained withinan intron of the ChAT gene, and some of its mRNAs are transcribed from the ChATpromoter (Bejanin et al., 1994; Erickson et al., 1994). The structure of the ChAT genehas been extensively studied and it has been shown to have several promoters inboth rat, mice and man (Chireux et al., 1994; Hahn et al., 1992; Ibáñez and Persson,1991; Kengaku et al., 1993; Misawa et al., 1992). We have previously isolated a2336 base pair (bp) long region from the most upstream promoter of the rat ChATgene that is able to confer cholinergic cell-specific expression of a reporter gene intransgenic mice (Lönnerberg et al., 1995). This transgene was also able to mimic thedevelopmental regulation of the endogenous ChAT gene and showed similarregulation in a lesion model (Lönnerberg et al., 1995).
Gene targeting in transgenic mice is a powerful approach to overexpress or inhibitspecific genes in a tissue-specific manner. A limitation of this approach, however, isthat it is often difficult to separate the effects observed in the adult from abnormalitiesin the developmental history of the animal caused by the genetic manipulation. Several system have been developed to overcome this problem and to allow theinvestigator to induce gene expression by exogenous administration of smalldifusible molecules. One such system was developed by Bujard and colleagues(Gossen et al., 1995), and it is based in the use of a chimeric transcritptional activatorwhich binding to specific DNA sequences is induced by tetracycline analogues. Thisstrategy has recently been applied to target tissue-specific regulatable transgeneexpression in several systems (Hennighausen et al., 1995; Kistner et al., 1996; Yu etal., 1996). We have now utilized this aspproach to produce a line of trnasgenic micethat selectively expresses the chimeric tetracycline-dependent transactivator rtTA incholinergic tissues.
Inducible and tissue-specific gene targeting to cholinergic neurons
The plasmids pUHD172-1neo, pUHD10-3 and pUHC13-6 were kindly provided by Dr. HermannBujard. pEH2.3TKrtTA expressing rtTA under control of the herpes simplex virus TK minimalpromoter and a cholinergic-specific regulatory region was constructed by inserting an EcoRI-BamHIfragment of the rtTA gene from pUHD172-1neo into the pBLEH2.3CAT plasmid. pBLEH2.3CATcontains the 2336 bp long EcoRI-HindIII regulatory region from the rat ChAT gene (Ibáñez andPersson, 1991; Lönnerberg et al., 1995) subcloned by blunt-end ligation into the SalI site ofpBLCAT (Luckow and Schütz, 1987). In the second construct tested, pEH2.3CMVrtTA, a CMV
minimal promoter was used instead of the TK promoter. This was done by inserting the 2336 bpEcoRI-HindIII regulatory region of the ChAT gene into the EcoRI site of pUHD10-3.
FR3T3 fibroblasts and the cholinergic SN6 cells were grown in DMEM with 10% of Fetal Calf Serumsupplemented with 2 mM glutamine, 100 U/ml penicillin, and 100 mg/ml streptomycin (all GibcoBRL) at 37oC in a 5% CO2 atmosphere. Cells grown to approximately 70% confluency weretransfected according to the calcium phosphate procedure. Doxycycline was added to the mediumimmediately after transfection. Forty eight hours later, the cells were harvested in lysis buffercontaining 0.1 M potassium phosphate buffer pH 7.7, 1 % Triton X-100, 15 % glycerol and 2mMdithiotreithol. Luciferase assays were performed as previously described (de Wet et al., 1987).
A 4.7 kb fragment containing the 2336 bp EcoRI-HindIII fragment from the regulatory region of therat ChAT gene followed by the TK minimal promoter, the rtTA gene and polyadenylationsequences was excised from pEH2.3TKrtTA, purified on an agarose gel, and injected in pronucleiof fertilized mouse eggs to generate transgenic mice as previously described (Nilsson and Lendahl,1993) (CBA x C57BL6)F1 mice were used as embryo donors, stud males, and pseudopregnantfemales. C57BL6 mature females were used for breeding. Preparation of high molecular weightDNA from tail biopsies was performed as previously described (Nilsson and Lendahl, 1993).
Tissue preparation, RNA extraction and RT-PCR analysis.
All tissues were immediately frozen on dry ice after dissection and stored at-70oC until analysis. RNA was extracted by the acid-phenol method (Chomczynski and Sacci, 1987)and polyadenylated RNA isolated with Dynabeads (Dynal). The expression of rtTA mRNA wasdetermined by reverse transcriptase-polymerase chain reaction (RT-PCR). 1 mg of RNA was reversetranscribed for 30 min at 60°C, then amplified with 5 cycles of 94°C for 45 s, 55°C for 1 min, 60°C for1 min, and 18 cycles of 94°C for 45 s and 60°C for 1 min (AmpliTaq EZ rTth RNA PCR kit, Perkin-Elmer). The primers used were 5’-GCCGCCATTATTACGACAAGC-3’ and 5’-GTCCTCGCCGTCTAAGTGGAG-3’. The products were run on an agarose gel, and photographedon a UV light table.
We have previously characterized a fragment from the rat choline acetyltransferasegene regulatory region that directs tissue-specific expression of a reporter gene in cell
Inducible and tissue-specific gene targeting to cholinergic neurons
lines and transgenic mice (Ibáñez and Persson, 1991; Lönnerberg et al., 1995;Lönnerberg et al., 1996). With the aim of obtaining reversible and regulatableexpression of genes targeted to cholinergic cells, we have combined the specificity ofthe enhancer region of the ChAT gene with the tetracycline inducible promoter system(Gossen et al., 1995).
Figure 1. Induction of luciferase activity by doxycycline in transfected cholinergic cells. Cholinergic SN6 cells and FR3T3 fibroblasts were transiently transfected with pUHC13-6 alone (-) and together (+) with (A) pEH2.3CMVrtTA or (B) pEH2.3TKrtTA. 2.5 mg of each plasmid were used for a 35 mm well. Doxycycline was added to indicated wells immediately after transfection at a concentration of 1 mg/ml. Cells were harvested after 48 hours incubation followed by assay of luciferase activity.
Cholinergic cell-type specific activation of a reporter gene by doxycyclin.
Two different promoters were tested for their efficiency at directing rtTA expression. Plasmids expressing rtTA under control of the 2336 bp cholinergic-specific regulatoryregion followed by the CMV (plasmid pEH2.3CMVrtTA) or TK (plasmidpEH2.3TKrtTA) promoters were transiently co-transfected together with a plasmidexpressing luciferase under control of heptamer tetop rtTA binding sites and the TK
Inducible and tissue-specific gene targeting to cholinergic neurons
minimal promoter (plasmid pUHC13-6) into the cholinergic neuroblastoma cell lineSN6 or into the rat fibroblast line FR3T3. For each plasmid pair, 1 mg/ml ofdoxycycline was added to one of two duplicate wells immediately after transfection. Forty eight hours later, the cells were collected and processed for luciferase assays. Doxycycline induced 7- and 13-fold activation of the reporter gene in cholinergic SN6cells that carried the pEH2.3CMVrtTA and pEH2.3TKrtTA constructs, respectively. Neither promoter showed any induction in non-cholinergic FR3T3 fibroblast cells (Fig. 1). The CMV promoter exhibited higher absolute expression levels than the TKpromoter when induced by doxycycline in SN6 cells, but displayed a significantbackground luciferase activity in the absence of doxycycline. No measurablebackground was detected with the pEH2.3TKrtTA construct (Fig. 1). Thus, cholinergic-specific inducibility was higher with the TK promoter construct, even though thispromoter had weaker overall activity than CMV.
Figure 2. Dose-dependent induction of luciferase activity by doxycycline in transfected SN6 cells. SN6 cells were transfected with 2.5 m g each of pUHC13-6 and pEH2.3TKrtTA. Immediately after transfection the indicated concentrations of doxycycline were added to the medium. Cells were grown for 48 hours, harvested, and processed for luciferase assays. Values shown represent mean +/- standard error (n=3).
Dose-dependent induction of the luciferase reporter gene
SN6 cells were transiently co-transfected with equal amounts of pEH2.3TKrtTA andpUHC13-6, and grown in the presence of various concentrations of doxycycline. Cellextracts were made after 48 hours, and luciferase enzyme activity was measured. Adose-dependent induction of the reporter gene was observed; maximal effects wereseen at a doxycycline concentration of 3000 ng/ml (Fig. 2).
Targeted expression of rtTA in cholinergic cells of transgenic mice
Inducible and tissue-specific gene targeting to cholinergic neurons
Having demonstrated the cholinergic-specificity and tetracycline-inducibility ofpEH2.3TKrtTA and pEH2.3CMVrtTA constructs in vitro, we set out to generate linesof transgenic mice with targeted expression of rtTA in cholinergic cells. Siince our goalwas to develop a system for reversible and regulatable gene targeting with minimaldevelopmental effects, we chose the construct based on the TK promoter as this isthe one that showed higher inducibility with neglible background activity, despite itsoverall lower activity compared to the CMV construct. The pEH2.3TKrtTA constructhas the rtTA coding region replacing the chloramphenicol acetyltransferase reportergene used in our earlier study of cholinergic-specific gene targeting in transgenic mice(Lönnerberg et al., 1995), and it was therefore expected to behave in a similarfashion.
Figure 3. Targeted expression of rtTA in cholinergic cells of transgenic mice. Total RNA was extracted from various tissues of an adult male of transgenic line 4, and RT-PCR was performed on 1 mg of total RNA for 31 cycles. The amplified products were run on an agarose gel, stained with ethidium bromide and photographed on a UV light table. No additional bands were detected after further 8 cycles of amplification.
Several lines of transgenic mice were obtained carrying the EH2.3TKrtTA transgene. Eight adult males each from a different line were sacrificed and various tissuesdissected for RNA extraction. Expression of rtTA mRNA was investigated byanalyziing alliquots of a RT-PCR reaction taken at different times of amplification. rtTAmRNA expression was detected in the spinal cord, which is rich in cholinergicmotorneuron cell bodies, in most of the lines (data not shown). One animal (transgenicline 4), expressing the highest level of rtTA mRNA in the spinal cord, was selected forfurther analysis. Fig. 3 shows the result of the survey of several tissues from thisanimal after 31 cycles of amplification and demonstrates the predominant expressionof rtTA mRNA in the spinal cord. Similar to several transgenic lines generated in ourprevious study (Lönnerberg et al., 1995), line 4 also showed transgene expressionin the cerebellum, despite no cholinergic cell bodies have been reported in thisregion. Further studies will be required to establish the cellular origin of rtTAexpression in this line using reporter genes suitable to histological analysis such as theE. coli gene product beta-galactosidase.
Inducible and tissue-specific gene targeting to cholinergic neurons
In conclusion, we have documented in vitro in cell culture experiments a system forregulatable cholinergic cell-specific expression of a reporter gene. We havefurthermore established several transgenic mice lines, one of which showspredominant expression of the rtTA transactivator in the spinal cord, which is therichest source of cholinergic cell bodies in the nervous system. These mice will b euseful tools for developmental and functional studies on cholinergic neurons. Forexample, targeted and regulatable expression of the CRE recombinase tocholinergic neurons would allow for specific gene-deletion in cholinergic cells. Thefunction of several important genes, such as neurotrophic factor receptors TrkA, TrkB,TrkC and c-Ret, may thus be studied in adult cholinergic neurons. Targeted, dose-dependent and reversible overexpression of neurotrophic factors in cholinergicneurons will also be possible using these mice. This should allow the investigation ofacute effects of neurotrophic factors in brain repair and learning without the complicationof developmental effects. Finally, this apporach may also be useful to reversible anddose-dependent targeting to the cholinergic system of specific toxins. This mayprove a novel and elegant approach to generate animal models of cholinergicdegeneration, which could be very useful to study the role of the cholinergic system incognition and behaviour, as well as to model neurodegenerative diseases of thecholinergic system, such as amyotrophic lateral sclerosis (ALS) and Alzheimer’sdisease.
We thank H. Bujard and M. Gossen for their kind gift of plasmid constructs for thetetracycline system. We also thank Ann-Sofie Nilsson and Annika Ahlsén for technicalassistance, and Lotta Johansson for secretarial help. This work was funded by grantsfrom the Swedish Medical Research Council, Centrala Försökdjursnämnden, LarsHiertas Minne and the Karolinska Institute.
Arvidsson, U., Riedl, M., Elde, R., and Meister, B. (1997). Vesicular acetylcholine transporter(VAChT) protein: A novel and unique marker for choliinergic neurons in the central and peripheralnervous systems. J Comp Neurol 378, 454-467.
Bejanin, S., Cervini, R., Mallet, J., and Berrard, S. (1994). A unique gene organization for twocholinergic markers, choline acetyltransferase and a putative transporter of acetylcholine. J BiolChem 269, 21944-21947.
Chireux, M., Raynal, J. F., Le Van Thai, A., Cadas, H., Bernard, C., Martinou, I., Martinou, J.-C., andWeber, M. B. (1994). Multiple promoters of human choline acetyltransferase and aromatic L-aminoacid decarboxylase genes. J Physiol Paris 88, 215-227.
Inducible and tissue-specific gene targeting to cholinergic neurons
Chomczynski, P., and Sacci, N. (1987). Single-step method of RNA isolation by guanidiniumthiocyanate-phenol-chloroform extraction. Anal Biochem 162, 156-159.
de Wet, J. R., Wood, K. V., DeLuca, M., Helinski, D. R., and Subramani, S. (1987). Firefly luciferasegene: structure and expression in mammalian cells. Mol Cell Biol 7, 725-737.
Erickson, J. D., Varoqui, H., Schäfer, M. K.-H., Modi, W., Diebler, M.-F., Weihe, E., Rand, J., Eiden,L. E., Bonner, T. I., and Usdin, T. B. (1994). Functional identification of a vesicular acetylcholinetransporter and its expression from a "cholinergic" gene locus. J Biol Chem 269, 21929-21932.
Gossen, M., Freundlieb, S., Bender, G., Müller, G., Hillen, W., and Bujard, H. (1995). Transcriptionalactivation by tetracyclines in mammalian cells. Science 268, 1766-1769.
Hahn, M., Hahn, S., Stone, D., and Joh, T. (1992). Cloning of the rat gene encoding cholineacetyltransferase, a cholinergic neuron-specific marker. Proc Natl Acad Sci USA 89, 4387-4391.
Hennighausen, L., Wall, R. J., Tillmann, U., Li, M., and Furth, P. A. (1995). Conditional geneexpression in secretory tissues and skin of transgenic mice using the MMTV-LTR and thetertracyclin responsive system. J Cellular Biochem 59, 463-472.
Ibáñez, C. F., and Persson, H. (1991). Localization of sequences determining cell type specificityand NGF responsiveness in the promoter region of the rat choline acetyltransferase gene. Eur JNeurosci 3, 1309-1315.
Kengaku, M., Misawa, H., and Deguchi, T. (1993). Multiple mRNA species of cholineacetyltransferase from rat spinal cord. Brain Res Mol Brain Res 18, 71-6.
Kistner, A., Gossen, M., Zimmermann, F., Jerecic, J., Ullmer, C., Lübbert, H., and Bujard, H. (1996). Doxycycline-mediated quantitative and tissue-specific control of gene expression in transgenicmice. Proc Natl Acad Sci USA 93, 10933-10938.
Luckow, B., and Schütz, G. (1987). CAT constructions with multiple unique restriction sites for thefunctional analysis of eukaryotic promoters and regulatory elements. Nucleic Acids Res 15, 5490.
Lönnerberg, P., Lendahl, U., Funakoshi, H., Ährlund-Richter, L., Persson, H., and Ibáñez, C. F. (1995). Regulatory region in choline acetyltransferase gene directs developmental and tissue-specific expression in transgenic mice. Proc Natl Acad Sci USA 92, 4046-4050.
Lönnerberg, P., Schoenherr, C. J., Anderson, D. J., and Ibáñez, C. F. (1996). Cell-specificregulation of the choline acetyltransferase gene: Role of the neuron-restrictive silencer elementand cholinergic-specific enhancer sequences. J. Biol. Chem. 271, 33358-33365.
Misawa, H., Ishii, K., and Deguchi, T. (1992). Gene expression of mouse choline acetyltransferase. Alternative splicing and identification of a highly active promoter region. J Biol Chem 267, 20392-20399.
Nilsson, E., and Lendahl, U. (1993). Transient expression of a human b-actin promoter/lacZ geneintroduced into mouse embryos correlates with a low degree of methylation. Mol Reprod Dev 34,149-157.
Yu, Z., Redfern, C. S., and Fishman, G. I. (1996). Conditional transgene expression in the heart. Circ Res 79, 691-697.
Pharmacogenetics of the 5-lipoxygenase biosyntheticpathway and variable clinical response to montelukastMichael Klotsmana, Timothy P. Yorkc, Sreekumar G. Pillaia,Cristina Vargas-Irwind, Sanjay S. Sharmaa, Edwin J.C.G. van den Oordband Wayne H. AndersonaObjective Interindividual clinical response to leukotrienemajority of patients with the wild-type alleles had only amodifiers is highly variable


 Inducible and tissue-specific gene targeting to cholinergic neurons
lines and transgenic mice (Ibáñez and Persson, 1991; Lönnerberg et al., 1995;Lönnerberg et al., 1996). With the aim of obtaining reversible and regulatableexpression of genes targeted to cholinergic cells, we have combined the specificity ofthe enhancer region of the ChAT gene with the tetracycline inducible promoter system(Gossen et al., 1995).
Inducible and tissue-specific gene targeting to cholinergic neurons
lines and transgenic mice (Ibáñez and Persson, 1991; Lönnerberg et al., 1995;Lönnerberg et al., 1996). With the aim of obtaining reversible and regulatableexpression of genes targeted to cholinergic cells, we have combined the specificity ofthe enhancer region of the ChAT gene with the tetracycline inducible promoter system(Gossen et al., 1995).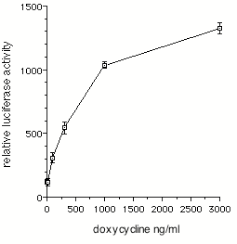 Inducible and tissue-specific gene targeting to cholinergic neurons
minimal promoter (plasmid pUHC13-6) into the cholinergic neuroblastoma cell lineSN6 or into the rat fibroblast line FR3T3. For each plasmid pair, 1 mg/ml ofdoxycycline was added to one of two duplicate wells immediately after transfection.
Inducible and tissue-specific gene targeting to cholinergic neurons
minimal promoter (plasmid pUHC13-6) into the cholinergic neuroblastoma cell lineSN6 or into the rat fibroblast line FR3T3. For each plasmid pair, 1 mg/ml ofdoxycycline was added to one of two duplicate wells immediately after transfection. Inducible and tissue-specific gene targeting to cholinergic neurons
Having demonstrated the cholinergic-specificity and tetracycline-inducibility ofpEH2.3TKrtTA and pEH2.3CMVrtTA constructs in vitro, we set out to generate linesof transgenic mice with targeted expression of rtTA in cholinergic cells. Siince our goalwas to develop a system for reversible and regulatable gene targeting with minimaldevelopmental effects, we chose the construct based on the TK promoter as this isthe one that showed higher inducibility with neglible background activity, despite itsoverall lower activity compared to the CMV construct. The pEH2.3TKrtTA constructhas the rtTA coding region replacing the chloramphenicol acetyltransferase reportergene used in our earlier study of cholinergic-specific gene targeting in transgenic mice(Lönnerberg et al., 1995), and it was therefore expected to behave in a similarfashion.
Inducible and tissue-specific gene targeting to cholinergic neurons
Having demonstrated the cholinergic-specificity and tetracycline-inducibility ofpEH2.3TKrtTA and pEH2.3CMVrtTA constructs in vitro, we set out to generate linesof transgenic mice with targeted expression of rtTA in cholinergic cells. Siince our goalwas to develop a system for reversible and regulatable gene targeting with minimaldevelopmental effects, we chose the construct based on the TK promoter as this isthe one that showed higher inducibility with neglible background activity, despite itsoverall lower activity compared to the CMV construct. The pEH2.3TKrtTA constructhas the rtTA coding region replacing the chloramphenicol acetyltransferase reportergene used in our earlier study of cholinergic-specific gene targeting in transgenic mice(Lönnerberg et al., 1995), and it was therefore expected to behave in a similarfashion.