Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Filestore.thermoscientific.com
JOURNAL OF VIROLOGY, Jan. 2001, p. 595–602
0022-538X/01/$04.00ϩ0 DOI: 10.1128/JVI.75.2.595–602.2001
Copyright 2001, American Society for Microbiology. All Rights Reserved.
Establishment of New Transmissible and Drug-Sensitive
Human Immunodeficiency Virus Type 1 Wild Types due to
Transmission of Nucleoside Analogue-Resistant Virus
ANTHONY DE RONDE,1* MAAIKE VAN DOOREN,1† LIA VAN DER HOEK,1 DENISE BOUWHUIS,1
ESTHER DE ROOIJ,1 BOB VAN GEMEN,2 ROB DE BOER,3 AND JAAP GOUDSMIT1
Department of Human Retrovirology, Academic Medical Center, University of Amsterdam,1105 AZ Amsterdam,1 Primagen, 1105 BA Amsterdam,2 and Theoretical Biology,Utrecht University, 3584 CH Utrecht,3 The Netherlands
Received 30 May 2000/Accepted 14 October 2000
Sequence analysis of human immunodeficiency virus type 1 (HIV-1) from 74 persons with acute infections identified eight strains with mutations in the reverse transcriptase (RT) gene at positions 41, 67, 68, 70, 215, and 219 associated with resistance to the nucleoside analogue zidovudine (AZT). Follow-up of the fate of these resistant HIV-1 strains in four newly infected individuals revealed that they were readily replaced by sensitive strains. The RT of the resistant viruses changed at amino acid 215 from tyrosine (Y) to aspartic acid (D) or serine (S), with asparagine (N) as a transient intermediate, indicating the establishment of new wild types. When we introduced these mutations and the original threonine (T)-containing wild type into infectious molecular clones and assessed their competitive advantage in vitro, the order of fitness was in accord with the in vivo observations: 215Y < 215D ؍ 215S ؍ 215T. As detected by real-time nucleic acid sequence-based amplification with two molecular beacons, the addition of AZT or stavudine (d4T) to the viral cultures favored the 215Y mutant in a dose-dependent manner. Our results illustrate that infection with nucleoside analogue- resistant HIV leads in newly infected individuals to mutants that are sensitive to nucleoside analogues, but only a single mutation removed from drug-resistant HIV. Such mutants were shown to be transmissible, stable, and prone to rapid selection for resistance to AZT or d4T as soon as antiretroviral therapy was administered. Monitoring of patients for the presence of new HIV-1 wild types with D, S, or N residues at position 215 may be warranted in order to estimate the threat to long-term efficacy of regimens including nucleoside analogues.
Since 1987, when antiviral drug therapy of human immuno-
HIV-1. The process does not necessarily lead back to a virus
deficiency virus type 1 (HIV-1) infection started with the use of
with a genotype identical to that of the original, drug-sensitive
zidovudine (AZT), drug-resistant viral mutants have rapidly
virus, but the phenotype (i.e., replicative capacity) will adapt to
emerged (3, 12). Due to the error-prone reverse transcriptase
be optimally fit in the new drug-free environment (7).
(RT), viral mutants are generated during every replication
Of 74 new HIV-1 infections that occurred in the period 1992
cycle (16). The mutational pathway is in essence the result of a
to 1999, 8 were caused by viruses with mutations at positions
stochastic process in which a single mutation appears more
associated with AZT resistance. After their transmission to
frequently than double or multiple mutations (13). A newly
new hosts, those with the resistance-conferring T215Y muta-
formed viral mutant that has a higher replicative capacity than
tion in the RT gene showed rapid evolution to better-replicat-
its parent will overgrow the parent. This process of selection
ing, more-fit viruses that acquired novel residues at amino acid
results in the appearance of mutants with the highest fitness or
215 of RT. The novel evolutionary pathway led to a viral
replicative capacity in their environment (7). When an antiret-
phenotype adapted to a drug-free environment and having a
roviral drug is present, mutants with the highest level of resis-
replicative capacity similar to that of the original wild type.
tance to the drug will have an advantage. As soon as antiviral
There was evidence that the new viruses were transmissible
therapy is stopped, the original virus, which is usually well
and rapidly converted by a single mutation to a drug-resistant
adapted to an environment without the drug, will win the
phenotype when suboptimal antiviral therapy was introduced.
competition with the resistant viruses (1). However, a different
situation occurs when a drug-resistant virus is transmitted to a
MATERIALS AND METHODS
previously uninfected and untreated person. Separated from
the original drug-sensitive virus and lacking this dominant
Study population. The Amsterdam cohort studies (ACS) have monitored a
competitor in the new drug-free environment, the drug-resis-
cohort of homosexual men and a cohort of intravenous drug users, with quarterlyscreening of participants for the presence of the HIV-1 antigen and antibodies.
tant virus will serve as a new starting point for the evolution of
New HIV-1 infections are identified by a seroconversion to the HIV-1 antigenand/or antibody with confirmation by Western blotting and HIV-1 viral loadassay. In addition, patients admitted to the clinic of the Academic Medical
* Corresponding author. Mailing address: Department of Human
Center (AMC) in Amsterdam with symptoms of suspected acute HIV-1 infection
Retrovirology, Academic Medical Center, University of Amsterdam,
have been examined. We studied a total of 74 new infections identified by ACS
Meibergdreef 15, 1105 AZ Amsterdam, The Netherlands. Phone:
and AMC between 1992 and 1999. Of these 74 new HIV-1 infections, 8 had RT
31.20.566.8571. Fax: 31.20.566.9080. E-mail: ronde@amc.uva.nl.
mutations that are associated with AZT-resistance (see below for the sequencing
† Present address: Primagen, 1105 BA Amsterdam, The Nether-
method). Mutations specifically associated with RT inhibitors other than AZT
(zalcitabine [ddC], didanosine, stavudine [d4T], lamivudine (3TC), nevirapine
and other nonnucleoside RT inhibitors) were not found. Of the 28 new HIV-1
ET47 (GAAGTGGGGATTCGACACACCAGACAAAAAAC; nt 3170 to
infections identified during or after 1996, the year of the introduction of protease
3209; sense; codon 215 is underlined) and ET48 (GTTTTTTGTCTGGTGTGT
inhibitors, none involved viruses that had mutations that confer resistance to
CGAATCCCCACTTC; nt 3209 to 3170; complementary) were used to intro-
protease inhibitors. Sequences of the 5Ј end of the RT gene with mutations
duce the 215 aspartic acid (GAC codon) mutation; ET49 (GAAGTGGGGAT
associated with AZT resistance were deposited in GenBank (see below for
TCTCCACACCAGACAAAAAAC; sense) and ET50 (GTTTTTTGTCTGGT
GTGGAGAATCCCCACTTC; complementary) were used to introduce the 215
Viral load and CD4 counts. Viral load was determined by using the NucliSens
serine (TCC codon) mutation; ET51 (GAAGTGGGGATTCACCACACCAGA
assay of Organon Teknika BV (Boxtel, The Netherlands). CD4ϩ cells in blood
CAAAAAAC; sense) and ET52 (GTTTTTTGTCTGGTGTGGTGAATCCCC
were counted by standard flow cytometry using a FACScan flow cytometer
ACTTC; complementary) were used to introduce the 215 threonine (ACC co-
(Becton Dickinson, San Jose, Calif.) and commercially available monoclonal
don) wild-type mutation. Sequencing was performed to verify that the exchanged
BstXI fragment contained the introduced mutations. By this procedure, we
Screening for resistance-conferring mutations. Viral RNA was isolated from
created plasmids p4-D, p4-S, and p4-T encoding the indicated (one-letter code)
200 l of serum drawn from each of the 74 subjects at the first point of sero-
conversion to the HIV-1 antigen or antibody by using the method described by
At week 154, viral RNA was isolated from a sample drawn from patient 4, and
Boom et al. (2). Part of the HIV-1 pol region including the part of the RT gene
RT-PCR was performed as described above using primers ET42 and ET10; the
encoding the amino terminus was amplified by RT-PCR, essentially according to
product was cloned in the TA cloning vector pCRII (InVitrogen BV). Sequence
the protocol described by Nijhuis et al. (18). To facilitate accurate two-stranded
analysis showed that individual clones were representative of the quasispecies
sequence analysis of the amino terminus-encoding part of the RT gene, we
that contained an S68G change relative to the p4-Y clone. In addition, these
adapted the nested PCR in the Nijhuis protocol by amplifying two overlapping
clones had naturally occurring variations at amino acids 39 and 135 (T39A,
fragments instead of one. The 5Ј fragment was amplified using primer SP6-
I135T). The 3Ј end of one of these clones (p4-68G) starting from the SspI site
RT19new, ATTTAGGTGACACTATAGCACCTGTCAACATAATTGGAAG
(HXB-2 nt 3025) was replaced by the 3Ј end of p4-Y or p4-D creating p4-Y-68G
(SP6 sequence is in italics; HXB-2 nucleotides [nt] 2491 to 2512), and T7-B-SEQ,
and p4-D-68G, respectively. The infectious molecular clone in which RT genes
TAATACGACTCACTATAGGGAATATTGCTGGTGATCCTTTCCA (T7 se-
derived from patient 4 were recombined was pHIV-Lai, lacking its RT gene
quence is in italics; HXB-2 nt 3030 to 2006). The 3Ј fragment was amplified with
(pLAI-⌬RT). The RT gene deletion was obtained by PCR-mediated mutagen-
primers SP6-C-SEQ, ATTTAGGTGACACTATAGGTATACTGCATTTACCA
esis. Briefly, the deletion was constructed in a subclone of HIV-Lai containing
TACC (SP6 sequence is in italics; HXB-2 nt 2927 to 2947), and T7-ET10, TAA
the ApaI-NcoI fragment by using primers ET40 (TGACCGCGGAAAATTTA
TACGACTCACTATAGGGCTGCCAGTTCTAGCTCTGCTTC (T7 sequence
AAGTGCAAC; nt 2550 to 2532; antisense; SacII site underlined) and ET23
is in italics; HXB-2 nt 3462 to 3441). Both strands of the nested PCR fragments
(AGTCCGCGGAGAGCAATGGCTAGT; nt 4286 to 4300; sense; SacII site
were directly sequenced using the SP6 and T7 primer sequences. Sequencing was
underlined) and primers 5Ј and 3Ј of ApaI and NcoI sites, respectively. This
performed with Taq dye primers (Applied Biosystems, Foster City, Calif.) and
created molecular clone pLAI-⌬RT with a SacII site replacing the RT gene.
the ThermoSequenase fluorescence-labeled primer cycle sequencing kit (Amer-
Recombinant viruses with a recipient-derived RT gene. To obtain recombinant
sham International, Little Chalfont, England). The sequence products were
viruses, C33A cells (human cervix carcinoma cell line) were seeded in 24-well
analyzed on an automatic sequencer (Applied Biosystems DNA sequencer
plates and grown to 80% confluence in 1 ml of RPMI 1640 medium (Life
model 373A stretch or 377). The sequence of the RT gene was screened at
Technologies, Breda, The Netherlands) supplemented with 10% fetal calf serum
positions associated with drug resistance as described by Hirsch et al. (10). For
and antibiotics. Cotransfection of 300 ng of pLAI-⌬RT and 200 ng of the purified
new infections caused by viruses with mutations at positions associated with AZT
RT gene-containing EcoRI fragment of p4-Y, -D, -S, -T, -Y-68G, or -D-68G was
resistance, serum samples drawn after the seroconversion point were analyzed
performed using the TFX-50 reagent protocol (Promega Benelux BV, Leiden,
for changes compared to the seroconversion sample. Mixtures of different viral
The Netherlands). One day after transfection, 200,000 MT2 cells per well were
RT sequences were quantified by assessing the ratio between the relevant peak
added, and C33A and MT2 cells were cocultured. After 2 days, the MT2 cells
areas of the signals of the corresponding nucleotides on the electropherogram
including medium were transferred to a 25-cm2 flask, with the addition of fresh
using the sequences of both strands. In cases identified during or after 1996, the
medium and MT2 cells. Cells were monitored for the formation of syncytia, and
year of introduction of protease inhibitors in The Netherlands, we sequenced the
the supernatant containing the recombinant virus was harvested after the spread
protease gene as well as the RT gene. The protease gene was amplified according
of syncytia throughout the culture. The virus was frozen into aliquots at Ϫ70°C.
to the procedure described by de Jong et al. (5). Nested PCR was performed by
The virus titer (50% tissue culture infective dose [TCID50]) was determined by
using primers SP6-A-SEQ, ATTTAGGTGACACTATAGGAGCCAACAGCCC
limiting-dilution titration on MT2 cells. Resistance to antiviral drugs was assayed
CACCAG (SP6 sequence is in italics; HXB-2 nt 2149 to 2167) and T7-A-SEQ,
TAATACGACTCACTATAGGGTAAAGAAGACAGTTACCG (T7 sequence is
Competition experiments were performed by mixing a total amount of 1,000
in italics; HXB-2 nt 2639 to 2621). Sequence analysis was performed as above for
TCID50 of the recombinant viruses in different ratios. These virus mixtures were
used to infect 2 ml of phytohemagglutinin-stimulated PBMC (2 ϫ 106 cells/ml),
Infectious molecular clones. Biological clones of the virus of patient 4 were
which were maintained in a six-well plate in RPMI 1640 medium supplemented
prepared by a limiting-dilution series of human peripheral blood mononuclear
with 10% fetal calf serum and interleukin-2 (6). Twice a week, cells were pas-
cells (PBMC) derived from a sample taken 1 month after seroconversion (14).
saged, and 10 l of the culture was transferred to 2 ml of a new culture of 2 ϫ
Viral RNA was isolated from the culture supernatants of individual biological
106 PBMC/ml. Similarly, 500,000 MT2 cells were infected with 1,000 TCID50 and
clones. The complete RT gene of the virus was reverse transcribed and amplified
maintained in 1 ml of RPMI medium supplemented with 10% fetal calf serum in
(5) but by using primers ET44 (GACATTTATCACAGCTGGCTAC; antisense;
a 24-well plate. When syncytia in the MT2 cells had formed, 5 l of the culture
nt 4338 to 4359 of HIVHXB2CG; GenBank accession no. K03455) and 5Ј-
was transferred to 1 ml of a new culture composed of 1 ml of medium containing
PROT-OUT (GAGCCAACAGCCCCACCAG; nt 2149 to 2167; sense) in the
500,000 MT2 cells. Competition experiments were performed in the absence or
RT reaction and first PCR and by using primers ET42 (CTGTTGACTCAGAT
presence of antiviral drugs (i.e., AZT, d4T, or ddC). At each passage, samples
TGGTTGCACTTTAAATTTTCC; nt 2517 to 2551; sense) and ET17 (AGGT
from the supernatant from which to isolate viral RNA for sequence analysis were
GGCAGGTTAAAATCAACTAGCCATGCCATTGCTCTCC; nt 4319 to 4285;
drawn. Viral RNA was isolated according to the method of Boom et al. (2). The
antisense) in the nested PCR. The PCR product was cloned in plasmid pCR-2.1
RNA was directly sequenced as described above. The ratios between the differ-
using a TA vector system (InVitrogen BV, Groningen, The Netherlands). Se-
ent recombinant viruses were determined from the direct sequence by assessing
quence analysis of the 5Ј end of the RT gene was performed, and a clone with the
the ratios between the relevant peak areas of the signals on the electropherogram
T215Y mutation (AZT resistant) was chosen. The insert was subcloned in the
corresponding to the nucleotides encoding amino acid 215 of RT. Alternatively,
EcoRI site of vector pUC19 to remove the flanking BstXI sites, creating p4-Y.
the ratios between the different viruses were determined by using real-time
PCR-mediated mutagenesis of amino acid 215 of RT was performed essentially
nucleic acid sequence-based amplification (NASBA) with molecular beacons, as
as described by de Jong et al. (5) except that primer ET10 (CTGCCAGTTCT
AGCTCTGCTTC; nt 3462 to 3441) and the sense mutagenesis primer for the
Quantification of mutant mixtures using real-time NASBA and molecular
first PCR were used, whereas primer ET07 (GGAAGTTCAATTAGGAATA
beacons. RNA was isolated from 100 l of culture supernatant according to the
CC; nt 2812 to 2832) and the antisense mutagenesis primer were used for the
method of Boom et al. (2). The nucleic acids were eluted in 50 l of water, and
second PCR. The two PCR products were combined, and a third PCR was
5 l of the eluate was used as input for amplification by NASBA. NASBA was
performed using primers ET10 and ET07. The resulting PCR product was
performed by using the basic kit of Organon Teknika supplemented with primers
digested with BstXI and cloned into a BstXI-digested p4-Y. Mutagenesis primers
215-P2 GACTTAGAAATAGGGCAGCA (HXB-2 nt 2705 to 2724) and 215-P1
TABLE 1. Example of the fitness calculation with
[H(t)/H(0)]/{ln[W(t)/W(0)] ϩ ␦t}, which is available at http://www-binf.bio.uu.nl
/ϳrdb/fitness.html. If the total viral loads and the percentages of wild-type virusat times 0 and t are known, the website equation can be used to compute the
change in the wild-type virus load, W(t)/W(0), and the change in the ratio of the
genotype frequencies, H(t)/H(0), to give s. H is undefined when the percentage
of mutant virus is estimated as 0 or 100%. Whenever possible, we therefore
ignored such data points. However, in a few instances (patients 2 and 4) no
alternative data points were available. In such instances, we took 1 or 99% for the
genotype frequency, which was a conceivable assumption (see Table 1 for the
example of patient 1). For the fitness calculations for the viral culture we took the
cumulative p24 value as the viral load, i.e., a p24 value of 1,000 pg/ml after a
1:100 dilution of a viral culture was 100,000 pg/ml, allowing us to keep a dilutionfactor of 1 in the formula on the website.
We calculated s over the interval between samples 1 and 6 (Table 1) (using 99
and 1% for the percentages of virus with Y at position 215 [215Y virus] in
The mixture contained 13% 215D virus.
samples 1 and 6, respectively); the fitness difference between 215Y and 215N was2.8%. Corresponding fitness differences were 2.3, 3.3, 2.6, 2.5, and 3.0% forintervals between samples 2 and 5, 2 and 4, 2 and 3, 3 and 4 (all of which used
AATTCTAATACGACTCACTATAGGGGTTCATAACCCATCCAAAGGAAT
detected percentages of 215Y virus), and 2 and 6 (which used 1% 215Y virus for
GGA (T7 sequence in italics; HXB-2 nt 2831 to 2806) and the molecular beacons
sample 6), respectively. The one-sample t test of the SSPS, version 8.0, statistical
(20) RT 215GAC, ccgactcTCGACACACCAGACAAAAAACgagtcgg (FAM,
package was used to calculate the mean and the 95% confidence interval. We
dabcyl), and RT 215TAC, ccgactcTCTACACACCAGACAAAAAACgagtcgg (6-
concluded that 215N had a relative fitness of 103% (95% confidence interval: 102
ROX, dabcyl) (stem sequence is in lowercase; HXB-2 nt 2772 to 2792; position
to 104%), with 215Y fitness ϭ 100%.
215 [underlined, boldface] is either a GAC [D] or TAC [Y] codon). The two
We calculated s over the interval between samples 1 and 4 for the virus culture
primers and two beacons were each added to a final concentration of 200 nM.
of the competition between 215Y and 215T viruses (Table 2); the fitness differ-
Fluorescence was measured in real time on a Fluoroscan Ascent (Labsystems Oy,
ence between 215Y and 215T was 5.1%. Corresponding fitness differences were
Helsinki, Finland). The assays were calibrated by mixing different quantities of in
5.4, 6.0, 4.7, and 4.9% for intervals between samples 2 and 4, 3 and 4, 1 and 3, and
vitro-synthesized RNA containing the GAC or TAC position 215 codon. To
2 and 3, respectively. We concluded that 215T had a relative fitness of 105%
synthesize the RNA, PCR fragments were generated from p4-D-68G and p4-Y-
(95% confidence interval: 104 to 106%), with 215Y fitness ϭ 100%.
68G by using primers AATTCTAATACGACTCACTATAGGG (T7 site located
Nucleotide sequence accession numbers. Accession numbers for viral RT gene
in the plasmid) and ET10 (HXB-2 nt 3462 to 3441). These PCR fragments were
sequences obtained in this study were as follows: patient 1 (H0671), AF265569;
used to produce RNA containing the GAC or TAC position 215 codon by using
patient 2 (I6056), AF265572; patient 3 (H0095), AF265570; patient 4 (I7052),
T7 RNA polymerase (Amersham Pharmacia Biotech Benelux, Roosendaal, The
AF265571; patient 5 (H0137), AF265568; patient 6 (I3234), AF265573; patient 7
Netherlands) in an in vitro transcription reaction. Under our reaction conditions
(M12690), AF265574; patient 8 (2202575), AF265567.
with 105 molecules of in vitro-synthesized RNA per reaction, the beacon specificfor the GAC codon did not react with the RNA containing the TAC codon and
vice versa. In vitro-synthesized RNA containing the GAC and TAC codons wasmixed at different percentages (0, 2, 4, 10, 33, 50, 67, 90, 96, 98, and 100%). In vivo evolution of viruses with a mutated RT gene in the
Repeated (n ϭ 5) experiments showed that quantification of the mixtures usingreal-time NASBA could be achieved until 4% of either the TAC or GAC variant
absence of an antiviral drug. Among 74 new HIV-1 infections
(see Fig. 4A and B). Below 4% of the variant, the variant could be observed but
occurring between 1992 and 1999, 8 were found to be due to
not quantified. Quantification of mixtures by real-time NASBA and direct se-
viruses that have mutations in the RT gene that are associated
quencing were compared for the competitions in the presence of AZT (see Fig.
with AZT resistance. At seroconversion, the corresponding
4C). The methods gave comparable percentages (Ϯ10%) of a variant mixture,provided that direct sequencing was performed using the ET dye primer on the
mutations in RT were M41L and T215Y (patients 1, 2, and 4);
ABI automatic sequencers and not the big-dye techniques on the ABI machines.
T215Y (patient 3); D67N, K70R, and T215F (patient 5);
However, when a variant was present in small amounts (less than 10%), quan-
D67N, K70R, and K219Q (patient 6); M41L and the T215D
tification by direct sequencing was no longer reliable due to background signals.
mutation, which involved an unusual amino acid not seen as-
Real-time NASBA was broader in dynamic range; it could reliably quantify a
sociated with AZT resistance (patient 7); and K70R (patient
variant at a level as low as 4% and could detect a variant at a level as low as 1%. Fitness calculation. Selection coefficients were calculated by a novel method
8). The K70R mutation in the virus infecting patient 8 could be
involving the estimation of the total viral replication during the observation
the result of a natural polymorphism, although in the absence
period (16a). The method can be used both for steady-state viral populations, to
of AZT the K70R variant rarely becomes the dominant variant
which most previously used methods apply (8), and for decreasing or expanding
in the viral quasispecies (17). The eight infections were mon-
viral populations, such as those observed, for example, in cell cultures. Briefly,the model is derived as follows. Because mutations in the RT are expected to
itored until antiviral therapy (Fig. 1). Infections 1 to 5, which
mainly influence the replication rate of a mutant virus and to hardly influence its
all were found at seroconversion to have an AZT resistance-
half-life, we write dW/dM ϭ rW Ϫ ␦W and dM/dt ϭ r(1 ϩ s)M Ϫ ␦M for the
conferring mutation at amino acid 215 of RT (either 215Y or
dynamics of wild-type virus W and mutant virus M. Here r is the (wild-type)replication rate (which may change over time [t]), 1/␦ is the generation time, ands is the classical coefficient of selection. Because this model combines the dy-
TABLE 2. Example of the fitness calculation with the virus culture
namics of productively infected cells and free virions, the generation time should
of the competition between 215Y and 215T viruses
be about 2 days (11). The same model has been used before for estimating s fromin vivo data with approximately steady-state viral loads (8). One conventionally
defines the frequency of the mutant genotype by the equation P ϭ M/(W ϩ M),
and hence 1 Ϫ p is the frequency of the wild-type virus. If r is a constant, one
writes the solutions W(t) ϭ W(0)e(r Ϫ ␦)t and M(t) ϭ M(0)e[r(1 ϩ s) Ϫ ␦]t so that s
is computed from the logarithms of the genotype ratios (H ϭ M/W ϭ p/[1 Ϫ p])
at time 0 and time t, i.e., s ϭ ln[H(t)/H(0)]/rt.
Thus, for estimating the relative fitness one generally needs to know the ratio
of the genotype frequencies and the total replication, rt. r may vary over time,however. This requires the integration of the replication rate over the experi-
ment. Fortunately, s can still be estimated by the simple formula (16a) s ϭ lnb Cumulative p24 in culture medium.
codon, with full replacement observed at week 20. At week 37,
we detected a mixture of the 215 GAC codon and the 215 TCC
codon (S), in which the relative abundances of the two fluctu-
ated until week 150, when the last sample was drawn before
therapy began. The changes to an aspartic acid and to a serine
at position 215 of RT required only a single mutation, from the
TAC codon to GAC and TCC codons, respectively. Analogous
to what was observed in patient 3, the RT gene of patient 4
showed a change at position 68. In this case, however, the
wild-type 68 AGC codon (serine) changed to a GGC codon
encoding glycine (68G). At week 120, wild-type 68S had been
completely replaced by 68G, which persisted thereafter. An
overview of the mutational pathways in the different patients is
The rate of RT evolution in these cases varied considerably.
In patient 1, replacement of 215Y by 215N and subsequently by
215D took place over a time period of more than 150 weeks.
Concurrent with the replacement of 215Y by 215N, the viral
FIG. 1. In vivo evolution of transmitted viruses with drug resis-
load increased from below 1,000 copies/ml to 10,000 copies/ml,
tance-conferring mutations in the absence of antiretroviral therapy.
and it increased further to 30,000 copies/ml when 215D be-
Indicated are the positions and amino acids (minor species in italics) in
came the dominant species. During this period, the CD4 T-cell
which the viruses differ from the wild type at positions known to be
involved in drug resistance (10). Our subjects were monitored until
counts remained stable and high at approximately 1,000/l. In
patient 2, replacement of 215Y by 215D occurred within less
than 40 weeks, with a viral load between 30,000 and 100,000
copies/ml and CD4 T-cell counts hovering between 400 and
215F) showed evolution in their RT genes. Since infections 1 to
600/l. In patient 3, replacement of 215Y by 215S occurred
4 could be monitored for at least 1 year before antiviral therapy
after 40 weeks; the viral load declined from 300,000 copies/ml
was given, the evolution of the RT gene, and in particular the
at seroconversion to 10,000 to 30,000 copies/ml from week 30
evolution at amino acid 215 of RT, was analyzed in these cases
to 55 and CD4 T cells remained at 200 to 400/l. In patient 4,
replacement of 215Y by 215D occurred within 25 weeks, with
In patient 1 at seroconversion, the RT of the infecting vi-
a viral load that declined from above 100,000 copies/ml at
ruses had the M41L and T215Y mutations conferring AZT
seroconversion to about 25,000 copies/ml at week 100 and with
resistance. While the codon at position 41 of the RT gene did
CD4 T-cell counts varying from approximately 300 to 600/l.
not change, amino acid codon 215 changed from TAC, encod-
In all three patients who had been infected with a virus in
ing tyrosine, to AAC, encoding asparagine. The 215N virus had
which the RT gene contained both the 41L and 215Y muta-
completely replaced 215Y within 100 weeks (2 years) after
tions, the dominant viral species that emerged contained an
seroconversion. Over the next 50 weeks, the AAC codon at
RT with the 41L and 215D mutations. Patient 7 was infected
position 215 was gradually replaced by a GAC codon encoding
with a virus containing an RT with these 41L and 215D mu-
aspartic acid, which remained the dominant species thereafter.
tations, indicating that a virus with these mutations is trans-
In patient 2 at seroconversion, the M41L and T215Y muta-
tions were likewise present and the mutants evolved as in
In vivo fitnesses of the newly emerged viruses with muta-
patient 1, but on a far smaller time scale. During the 36 weeks
tions in the RT gene. The increased replicative capacity, or
that patient 2 was monitored 215Y evolved to a mixture of
fitness, of the newly emerged viruses with changes in their RT
215N and 215D, in which 215D eventually became the domi-
genes was expressed relative to the replicative capacity of the
nant species, a process that took 150 weeks in patient 1.
215Y variant at seroconversion, which was arbitrarily set at
In patient 3, RT had only the T215Y mutation at serocon-
100%. The fitnesses of the variants newly formed in an envi-
version, with codon 215 evolving from TAC (215Y) to AGC,
ronment without an antiviral drug were calculated. In patient 1
encoding serine (215S), although at week 11 GAC (215D) was
the available serum samples allowed the calculation of the
transiently observed. Unlike the other mutations we studied,
fitness of the 215N variant, which was 103% (95% confidence
which involve a one-nucleotide change within the codon 215,
interval: 102 to 104%) of that of 215Y in an environment
the change from TAC to AGC involved a two-nucleotide
without an antiviral drug. By the same method, the fitness of
change, and no obvious intermediate was detected. However,
the 215S variant (patient 3) was calculated to be 106% (103 to
in this infection, the 215 AAC codon may have been transiently
109%) of that of 215Y and the fitness of the 215D variant was
formed as an intermediate to the 215 AGC codon encoding
calculated to be 107 (patient 2) or 110% (patient 4) (96 to
serine. A notable mutation was observed at codon 68 of the RT
124%) of that of the 215Y variant. By way of comparison, the
gene, which changed from wild-type AGC (serine) to a mixture
S68G change observed in the RT of patient 4 resulted in a
relative fitness of the 68G viruses of 101% compared to that of
In patient 4, RT had the M41L and T215Y mutations at
the 68S virus, which was set at 100%. To give an impression of
seroconversion. At week 3 after seroconversion, the 215 TAC
the competition between the new variants and the AZT-resis-
(Y) codon was already being replaced by the 215 GAC (D)
tant 215Y, we plotted the fractions (relative to 215Y) of the
FIG. 2. In vivo evolution at amino acid 215 of RT. (A) Relative abundances of viruses with the indicated amino acids at position 215 of RT
after transmission of AZT-resistant viruses. The evolution of transmitted viruses is shown in patients 1 to 4. All start with 215Y and evolve to 215D
(patients 1, 2, and 4) and/or 215S (patients 3, and 4), with 215N observed as an intermediate (patients 1 and 2). (B) In vivo mutational pathways
as observed (patients 1, 2, and 4) or hypothesized (patient 3) at amino acid 215 of RT after transmission of AZT-resistant viruses.
different variants in the viral population (on a log scale) against
type 215T was found also in the viruses of the donor of patient
the elapsed time after seroconversion (Fig. 3A).
4 before he received AZT and resistant viruses developed. The
In vitro fitnesses of viruses differing at amino acid 215 of
various RT genes were recombined into an HIV-1 Lai back-
RT. In vivo, if HIV-1 isolates found in two different samples
ground, creating viruses that differed only at amino acid 215 of
drawn from a patient show genetic differences, they differ usu-
RT. Competition between the resulting viruses was observed in
ally at multiple positions, of which only a few may significantly
PBMC in the absence of an antiviral drug. In that environment,
contribute to a change in replicative capacity. To examine the
the 215Y virus was the least fit virus. The relative fitnesses of
significance of RT amino acid 215 for replicative capacity,
all the viruses were calculated. The in vitro relative fitnesses of
viruses that differed only at this amino acid were constructed.
215D, 215S, and 215T compared to that of 215Y (100%) were
In the RT isolated from patient 4, containing the M41L and
104 (95% confidence interval: 102 to 106%), 107 (104 to
T215Y mutations, only the 215 position was changed by in vitro
109%), and 105% (104 to 106%), respectively (Fig. 3B). Com-
mutagenesis to either 215D, 215S, or to 215T, the wild-type
petition among the 215D and the 215S or 215T viruses showed
amino acid present before the introduction of AZT. The wild-
them to be equally fit, i.e., having a fitness difference of less
mutations at positions 68 (S68G), 39 (T39A), and 135 (I135T).
To examine whether the change of environment caused by the
therapy with d4T and ddC was involved in the selection of a
viral population with the 215D- or 215S-to-215Y mutation,
viruses that differed only at amino acid 215 of RT were studied.
The viruses tested had either 215D or 215Y with 68G (present
directly before and after therapy) or 68S (present directly after
transmission). By using competition experiments with MT2
cells, the replicative capacities of the viruses were compared at
increasing concentrations of AZT, d4T, or ddC. To facilitate
the analysis of the competition experiments, we developed a
real-time NASBA with two molecular beacons differing at one
nucleotide. The dynamic range of quantification and the
throughput of the real time NASBA are higher than those in
sequence analysis, which generally cannot quantify mixtures
with less than 10% of a variant. The molecular beacons, among
which the GAC beacon contained a FAM label and the TAC
beacon contained a ROX label, were able to discriminate vi-
ruses differing at the 215 codon of the RT gene (Fig. 4A). The
real-time NASBA was able to quantify mixtures containing as
little as 4% of either the GAC- or TAC-containing virus (Fig.
4B) and to detect the GAC- or TAC-containing virus in a
mixture in which they composed no more than 1%. Using the
real-time NASBA, the analysis of the competition experiments
showed that the background of the viruses with the 68S or 68G
FIG. 3. In vivo and in vitro relative fitnesses of viruses marked by
mutation (either those directly after transmission or those
their amino acids at position 215 of RT. (A) Fitness plot of viruses
present at therapy) did not influence the phenotype. However,
differing at amino acid 215 (N, S, and D), which was derived from the
the 215D-to-215Y change was crucial to obtain a replicative
observed replacement of 215Y by viral variant 215N, 215S, or 215D in
advantage in the environment with both AZT and d4T (Fig.
the four patients (two of them had a 215D). The data points corre-
spond to the observed viral mixtures in the patients (Fig. 2A). Shown
4C). The replicative advantage of a virus with a 215Y codon in
are the rates (weeks after seroconversion) at which and to what extent
the presence of AZT or d4T varied in a dose-dependent man-
(on a log scale) 215Y virus is replaced by a different variant. (B)
ner. The replicative advantage of 215Y virus could be ex-
Fitness plot of viruses differing only at amino acid 215 of RT, which
pressed as an average fitness difference of 5 and 11% in the
competed in PBMC in the absence of an antiretroviral drug. Shown are
the rates (number of days of culture) at which and to what extent (on
presence of 45 and 170 nM AZT, respectively (0.5 and 2 times,
a log scale) variants 215D (}), 215S (■), and 215T (F) replaced 215Y.
respectively, the 50% inhibitory concentration [IC50] of a sen-
When 215D was competed with 215T or 215S, 215D was found to be
sitive virus), and 2 and 28% in the presence of 3 and 12 M
d4T, respectively (0.5 and 2 times, respectively, the IC50 of a
sensitive virus), indicating that the replicative advantage of
than 1%. The order of in vitro fitness of the viruses differing
215Y increased with higher drug concentrations. At 1.5 and 4.5
only at amino acid 215 was 215Y Ͻ 215D ϭ 215S ϭ 215T,
M (0.5 and 1.5 times, respectively, the IC50 of a sensitive
corresponding to the in vivo-observed order.
virus), ddC did not appear to influence the relative replicative
Characterization of the viruses after antiviral therapy by
capacities of the viruses. In classical phenotypic resistance as-
using real-time NASBA. Patient 1 was not treated because his
says, the 215Y virus is Ͼ100-fold more resistant to AZT and
CD4 counts remained stable and high at approximately 1,000
2-fold more resistant to d4T than the wild type and sensitive to
CD4 cells/l. Patients 2 and 3 were treated with a combination
ddC, whereas the 215D and 215S viruses are sensitive to AZT,
of protease and RT inhibitors (indinavir plus ritonavir plus d4T
plus 3TC and indinavir plus d4T plus 3TC), and their viral
loads declined to below detection level (Ͻ400 copies/ml). Pa-
DISCUSSION
tient 4 was treated at 160 weeks after seroconversion with RT
inhibitors d4T and ddC. No change in the viral load was ob-
Our study indicates that between 1992 and 1999, 10 to 15%
served between week 154 (pretherapy) and week 170 (after
of new HIV-1 infections in Amsterdam, The Netherlands, were
therapy), indicative of the development of resistance to the
caused by HIV-1 strains with mutations in the RT gene at
antiviral drug. In this case, the only observed mutation in RT
positions that are associated with AZT resistance. We found
was at amino acid 215, which was changed from a mixture of
no mutations associated with resistance to other antiretroviral
215D and 215S to 215Y by a single mutation (GAC and/or
drugs, including the protease inhibitors introduced in 1996.
TCC to TAC). Phylogenetic analysis of the RT genes of the
When the HIV-1 strains with drug resistance-conferring mu-
viruses found before and after therapy showed that the post-
tations are transmitted, a new environment without antiviral
therapy 215Y virus was derived from viruses present before
drugs in a new host is encountered. After transmission of
therapy but not present at transmission. The newer viruses
usually a small amount of virus, the wild-type strains from
shared 215Y with the transmission virus but had, in addition,
which the drug-resistant strains were derived are lost from the
FIG. 4. In vitro competitions of 215Y and 215D viruses in the absence and presence of antiviral drugs as analyzed by using real-time NASBA.
(A) Representative experiment in which relative fluorescence units (RFU) are shown as a function of time when RNA mixtures (total of 105
molecules) of 215D and 215Y (ranging from a 25:1 to a 1:25 ratio) were assayed by real-time NASBA using GAC (FAM-labeled) and TAC
(ROX-labeled) molecular beacons indicative of 215D and 215Y, respectively. (B) Calibration curve derived from panel A. The log function of the
slope of the increase of fluorescence in time is proportional to the relative abundances of 215D and 215Y in a sample, which can be inferred from
the known mixtures. (C) Competition between 215Y and 215D viruses in the presence of increasing concentrations of d4T (left) or AZT (right).
Viruses containing RT with 215Y and 215D competed in MT2 cells in the presence of 0, 3, or 12 M d4T or 0, 45, or 170 nM AZT. The ratio
of 215Y/215D virus in the supernatant at passage was determined by using real-time NASBA.
(memory of the) quasispecies and thereby are excluded from
215S virus. Compared to the transmitted AZT-resistant 215Y
competition in the new environment (19). The separation of
virus the 215N viruses were 3% more fit, the 215S viruses were
individual viruses from the remainder of competing viruses by
6% more fit, and the 215D viruses were 7 to 10% more fit. Our
bottleneck transmission is reminiscent of the founding of an
calculations assumed a constant viral generation time; how-
island population and may lead to new directions in evolution
ever, variation among the respective infections could affect the
(15). In the new, drug-free host, viruses that have an increased
relative fitness values. In this respect, patient 1, with an initially
replicative capacity or fitness compared to that of their drug-
low viral load and slow replacement of the different variants, is
resistant parent will be selected. Although the replicative prop-
illustrative. Nevertheless, the order of viral fitness is 215-ty-
erties of the viruses we studied will most likely be determined
rosine Ͻ 215-asparagine Ͻ 215-aspartic acid ϭ 215-serine. In
by more than one gene, we focused on the resistance-confer-
vitro experiments in a drug-free environment showed that wild-
ring RT gene. We found that the drug-resistance-conferring
type viruses with 215-threonine were as fit as the 215D or 215S
RT amino acid 215Y in viruses changed after transmission, but
viruses and fitter than the 215Y viruses. Threonine (ACC co-
not to the wild-type threonine. Instead, novel amino acids at
don; 215T) can be formed by a two-step mutational process
position 215 were observed (see also reference 21). Single
from the TAC codon (215Y). Such a two-step mutant, gener-
mutations in the TAC codon (215Y) gave rise to AAC (aspar-
ated by stochastic mutational processes, will usually be formed
agine), GAC (aspartic acid), or TCC (serine). In addition, in
later than one-step mutants such as 215D (GAC) and 215S
one of the new infections an AGC codon (serine, 215S) was
(TCC). The principle of competitive exclusion could explain
formed by a two-step mutation. Selection occurred for the
why we observed no 215T viruses in the new infections started
fittest variant, the 215N virus being less fit than the 215D or
with AZT-resistant viruses, since an equally fit variant will not
replace an existing virus that already dominates the population
cation of nucleic acids. J. Clin. Microbiol. 28:495–503.
(4). In this group, the phenotype of a 215D or 215S virus will
3. Boucher, C. A., E. O’Sullivan, J. W. Mulder, C. Ramautarsing, P. Kellam, G. Darby, J. M. Lange, J. Goudsmit, and B. A. Larder. 1992. Ordered appear-
to a large extent be the new wild type. This is most clearly
ance of zidovudine resistance mutations during treatment of 18 human
shown in the virus of patient 7, in which the 41L and 215D
immunodeficiency virus-positive subjects. J. Infect. Dis. 165:105–110.
mutations appeared directly at seroconversion, strongly point-
4. Clarke, D. K., E. A. Duarte, S. F. Elena, A. Moya, E. Domingo, and J. Holland. 1994. The Red Queen reigns in the kingdom of RNA viruses. Proc.
ing at the transmissibility of the 215D virus.
Natl. Acad. Sci. USA 91:4821–4824.
When antiviral therapy is applied, the environment changes
5. de Jong, J. J., J. Goudsmit, V. V. Lukashov, M. E. Hillebrand, E. Baan, R.
once yet again, and under conditions of suboptimal therapy in
Huismans, S. A. Danner, J. H. ten Veen, F. de Wolf, and S. Jurriaans. 1999.
Insertion of two amino acids combined with changes in reverse transcriptase
which sufficient residual viral replication can occur, a new
containing tyrosine-215 of HIV-1 resistant to multiple nucleoside analogs.
selection for the most fit variant takes place. For patient 4, a
AIDS 13:75–80.
single mutation (GAC or TCC to TAC) within the codon of
6. de Ronde, A., B. Klaver, W. Keulen, L. Smit, and J. Goudsmit. 1992. Natural
HIV-1 NEF accelerates virus replication in primary human lymphocytes.
amino acid 215 of RT appeared sufficient to confer a decisive
Virology 188:391–395.
replicative advantage in the presence of d4T and ddC. By using
7. Domingo, E., and J. J. Holland. 1997. RNA virus mutations and fitness for
a newly developed real-time NASBA with a high throughput
survival. Annu. Rev. Microbiol. 51:151–178.
8. Goudsmit, J., A. de Ronde, E. de Rooij, and R. de Boer. 1997. Broad
and dynamic quantification range, in vitro competitions with
spectrum of in vivo fitness of human immunodeficiency virus type 1 sub-
increasing concentrations of AZT or d4T showed that the
populations differing at reverse transcriptase codons 41 and 215. J. Virol. 71:
change from 215D to 215Y by a single nucleotide mutation
9. Hertogs, K., M. P. de Bethune, V. Miller, T. Ivens, P. Schel, A. van Cau-
conferred the replicative advantage. Apparently, the AZT-re-
wenberge, E. C. van den Eynde, V. van Gerwen, H. Azijn, M. van Houtte, F.
sistant genotype of the virus of patient 4 is also advantageous
Peeters, S. Staszewski, M. Conant, S. Bloor, S. Kemp, B. Larder, and R. Pauwels. 1998. A rapid method for simultaneous detection of phenotypic
in the presence of d4T. This finding shows that although the
resistance to inhibitors of protease and reverse transcriptase in recombinant
transmitted AZT-resistant viruses that have adapted to the
human immunodeficiency virus type 1 isolates from patients treated with
drug-free environment closely resemble the 215T wild type in
antiretroviral drugs. Antimicrob. Agents Chemother. 42:269–276.
10. Hirsch, M. S., B. Conway, R. T. D’Aquila, V. A. Johnson, F. Brun-Vezinet, B.
their phenotypes, their genotype retains many characteristics
Clotet, L. M. Demeter, S. M. Hammer, D. M. Jacobsen, D. R. Kuritzkes, C.
of the transmitted AZT-resistant 215Y virus. Loveday, J. W. Mellors, S. Vella, and D. D. Richman. 1998. Antiretroviral
Our observations confirm the high plasticity of the genome
drug resistance testing in adults with HIV infection: implications for clinical
management. International AIDS Society—USA Panel. JAMA 279:1984–
of HIV-1 and in particular of the RT gene. Transmission from
a treated host to an untreated host immediately results in
11. Ho, D. D., A. U. Neumann, A. S. Perelson, W. Chen, J. M. Leonard, and M.
adaptation of the virus to its new, drug-free environment. Such
Markowitz. 1995. Rapid turnover of plasma virions and CD4 lymphocytes in
HIV-1 infection. Nature 373:123–126.
adaptation becomes manifest as a fixation of a 215D or 215S
12. Kellam, P., C. A. Boucher, and B. A. Larder. 1992. Fifth mutation in human
mutation instead of reversion to the wild type of the pre-
immunodeficiency virus type 1 reverse transcriptase contributes to the de-
velopment of high-level resistance to zidovudine. Proc. Natl. Acad. Sci. USA
therapy era: 215T. These new wild types share AZT sensitivity
89:1934–1938.
with the old wild type, 215T. However, the new wild types are
13. Keulen, W., C. Boucher, and B. Berkhout. 1996. Nucleotide substitution
only one step away from 215Y and 215Y is therefore expected
patterns can predict the requirements for drug-resistance of HIV-1 proteins.
Antivir. Res. 31:45–57.
to arise rapidly upon administration of antiretroviral therapy
14. Koot, M., A. B. van’t Wout, N. A. Kootstra, R. E. de Goede, M. Tersmette,
when therapy is suboptimal. As we have observed, the newly
and H. Schuitemaker. 1996. Relation between changes in cellular load,
established wild types can be transmitted. The monitoring of
evolution of viral phenotype, and the clonal composition of virus populations
in the course of human immunodeficiency virus type 1 infection. J. Infect.
new wild types with D, S, or N at position 215 may thus be
Dis. 173:349–354.
warranted in order to estimate the threat to long-term efficacy
15. Losos, J. B., T. R. Jackman, A. Larson, K. Queiroz, and L. Rodriguez-
of a therapy regimen that includes nucleoside analogues. Schettino. 1998. Contingency and determinism in replicated adaptive radi-
ations of island lizards. Science 279:2115–2118.
16. Mansky, L. M., and H. M. Temin. 1995. Lower in vivo mutation rate of
human immunodeficiency virus type 1 than that predicted from the fidelity of
ACKNOWLEDGMENTS
purified reverse transcriptase. J. Virol. 69:5087–5094.
We thank M. Bakker and S. Jurriaans for providing serum samples;
16a.Maree, A. F. M., W. Keulen, C. A. B. Boucher, and R. J. De Boer. 2000.
K. Lindenberg and R. Coutinho of the Amsterdam Municipal Health
Estimating relative fitness in viral competition experiments. J. Virol. 74:
Service for patient data; V. Benes and H. Voss of EMBL for initial
17. Na´jera, I., A´. Holguı´n, M. E. Quin˜ones-Mateu, M. A´. Mun˜oz-Ferna´ndez, R.
help with sequencing; B. Hemmelder and J. Maas for technical assis-
Na´jera, C. Lo´pez-Galı´ndez, and E. Domingo. 1995. pol gene quasispecies of
tance; L. Phillips for editorial assistance; B. Berkhout, M. Cornelissen,
human immunodeficiency virus: mutations associated with drug resistance in
and V. Lukashov for discussion and ideas; and the participants of the
virus from patients undergoing no drug therapy. J. Virol. 69:23–31.
ACS for their cooperation over many years.
18. Nijhuis, M., C. A. Boucher, and R. Schuurman. 1995. Sensitive procedure for
This research was supported by grants 1311 and 1317 of the Dutch
the amplification of HIV-1 RNA using a combined reverse-transcription and
amplification reaction. BioTechniques 19:178–180.
19. Ruiz-Jarabo, C. M., A. Arias, E. Baranowski, C. Escarmis, and E. Domingo.
2000. Memory in viral quasispecies. J. Virol. 74:3543–3547. REFERENCES
20. Tyagi, S., D. P. Bratu, and F. R. Kramer. 1998. Multicolor molecular beacons
1. Albert, J., J. Wahlberg, J. Lundeberg, S. Cox, E. Sandstrom, B. Wahren, and
for allele discrimination. Nat. Biotechnol. 16:49–53. M. Uhlen. 1992. Persistence of azidothymidine-resistant human immunode-
21. Yerly, S., A. Rakik, S. K. De Loes, B. Hirschel, D. Descamps, F. Brun-
ficiency virus type 1 RNA genotypes in posttreatment sera. J. Virol. 66:5627– Vezinet, and L. Perrin. 1998. Switch to unusual amino acids at codon 215 of
the human immunodeficiency virus type 1 reverse transcriptase gene in
2. Boom, R., C. J. Sol, M. M. Salimans, C. L. Jansen, P. M. Wertheim-van
seroconvertors infected with zidovudine-resistant variants. J. Virol. 72:3520– Dillen, and J. van der Noordaa. 1990. Rapid and simple method for purifi-
Tadiran Medium Power Lithium Organic Cell Model TLM-1520MP This data sheet describes the mechanical design and performance of Tadiran medium power lithium organic cell model TLM-1520MP. 2. Characteristics (for batteries stored at RT for 1 year or less) 2.2.2. Closed Circuit Voltage (at 0.1 sec) at 0.25 A load Discharge capacity at 12 mA @ RT to 2.8 V Discharge capacity at 125 mA
OGA MEDIUM (OXYTETRACYCLINE GLUCOSE AGAR BASE) CAT Nº: 1527 For the enumeration and isolation of yeasts and molds in food samples FORMULA IN g/l Final pH 6.5 ± 0.2 at 25ºC PREPARATION Suspend 15 grams of the medium in 500ml of distil ed water. Mix well and dissolve by heating with frequent agitation. Boil for one minute until complete dissolution. Sterilize in autoclav
 codon, with full replacement observed at week 20. At week 37,
we detected a mixture of the 215 GAC codon and the 215 TCC
codon (S), in which the relative abundances of the two fluctu-
ated until week 150, when the last sample was drawn before
therapy began. The changes to an aspartic acid and to a serine
at position 215 of RT required only a single mutation, from the
TAC codon to GAC and TCC codons, respectively. Analogous
to what was observed in patient 3, the RT gene of patient 4
showed a change at position 68. In this case, however, the
wild-type 68 AGC codon (serine) changed to a GGC codon
encoding glycine (68G). At week 120, wild-type 68S had been
completely replaced by 68G, which persisted thereafter. An
overview of the mutational pathways in the different patients is
The rate of RT evolution in these cases varied considerably.
codon, with full replacement observed at week 20. At week 37,
we detected a mixture of the 215 GAC codon and the 215 TCC
codon (S), in which the relative abundances of the two fluctu-
ated until week 150, when the last sample was drawn before
therapy began. The changes to an aspartic acid and to a serine
at position 215 of RT required only a single mutation, from the
TAC codon to GAC and TCC codons, respectively. Analogous
to what was observed in patient 3, the RT gene of patient 4
showed a change at position 68. In this case, however, the
wild-type 68 AGC codon (serine) changed to a GGC codon
encoding glycine (68G). At week 120, wild-type 68S had been
completely replaced by 68G, which persisted thereafter. An
overview of the mutational pathways in the different patients is
The rate of RT evolution in these cases varied considerably.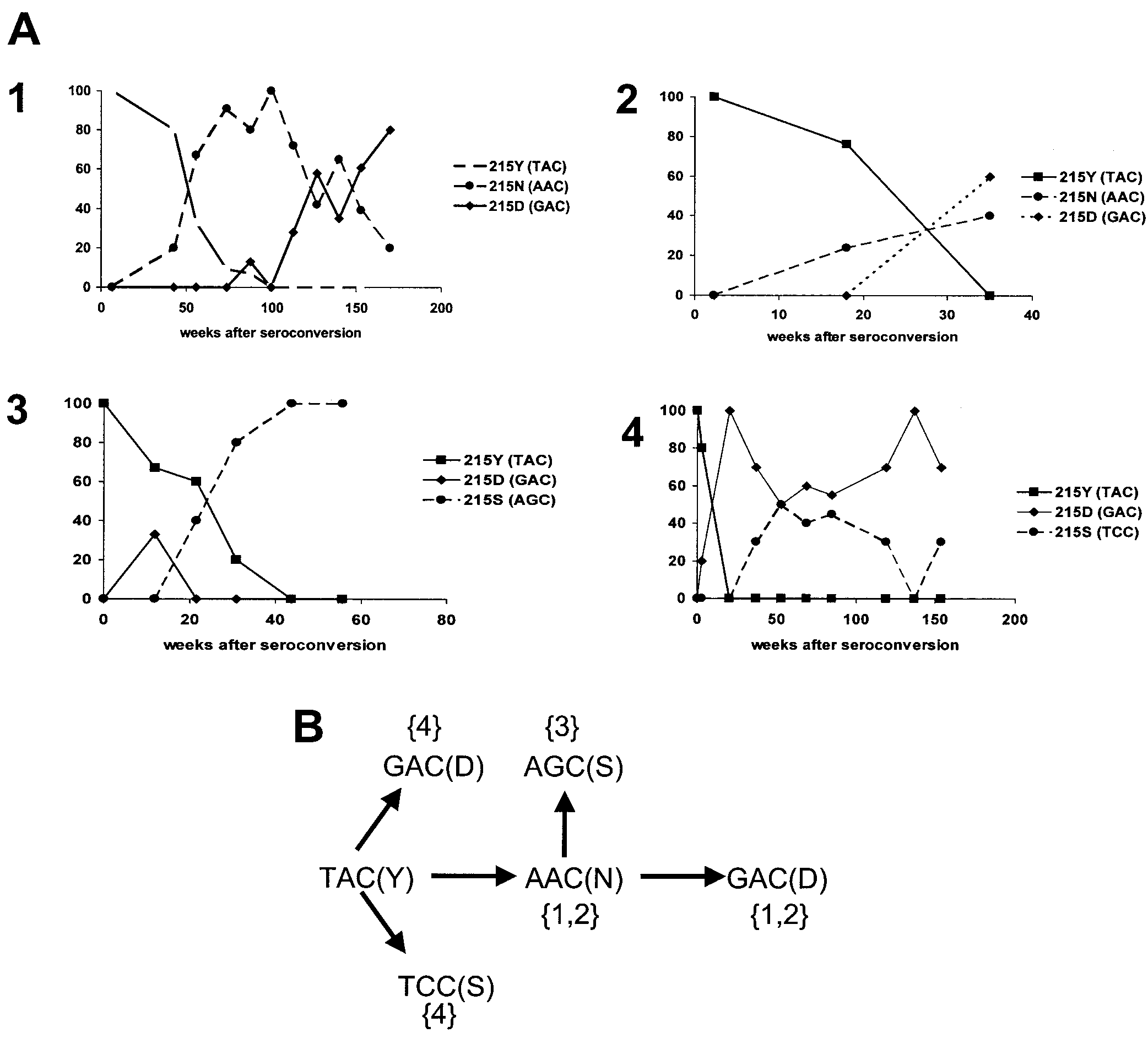 FIG. 2. In vivo evolution at amino acid 215 of RT. (A) Relative abundances of viruses with the indicated amino acids at position 215 of RT
after transmission of AZT-resistant viruses. The evolution of transmitted viruses is shown in patients 1 to 4. All start with 215Y and evolve to 215D
(patients 1, 2, and 4) and/or 215S (patients 3, and 4), with 215N observed as an intermediate (patients 1 and 2). (B) In vivo mutational pathways
as observed (patients 1, 2, and 4) or hypothesized (patient 3) at amino acid 215 of RT after transmission of AZT-resistant viruses.
FIG. 2. In vivo evolution at amino acid 215 of RT. (A) Relative abundances of viruses with the indicated amino acids at position 215 of RT
after transmission of AZT-resistant viruses. The evolution of transmitted viruses is shown in patients 1 to 4. All start with 215Y and evolve to 215D
(patients 1, 2, and 4) and/or 215S (patients 3, and 4), with 215N observed as an intermediate (patients 1 and 2). (B) In vivo mutational pathways
as observed (patients 1, 2, and 4) or hypothesized (patient 3) at amino acid 215 of RT after transmission of AZT-resistant viruses.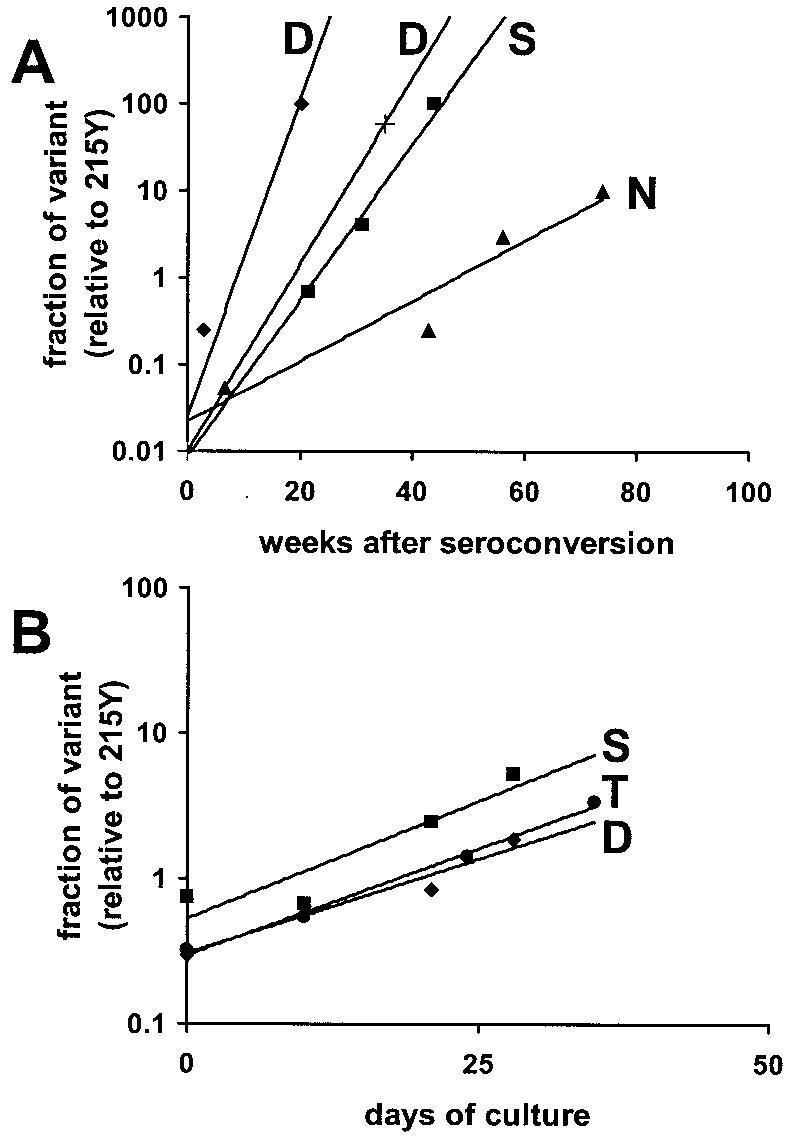 mutations at positions 68 (S68G), 39 (T39A), and 135 (I135T).
mutations at positions 68 (S68G), 39 (T39A), and 135 (I135T).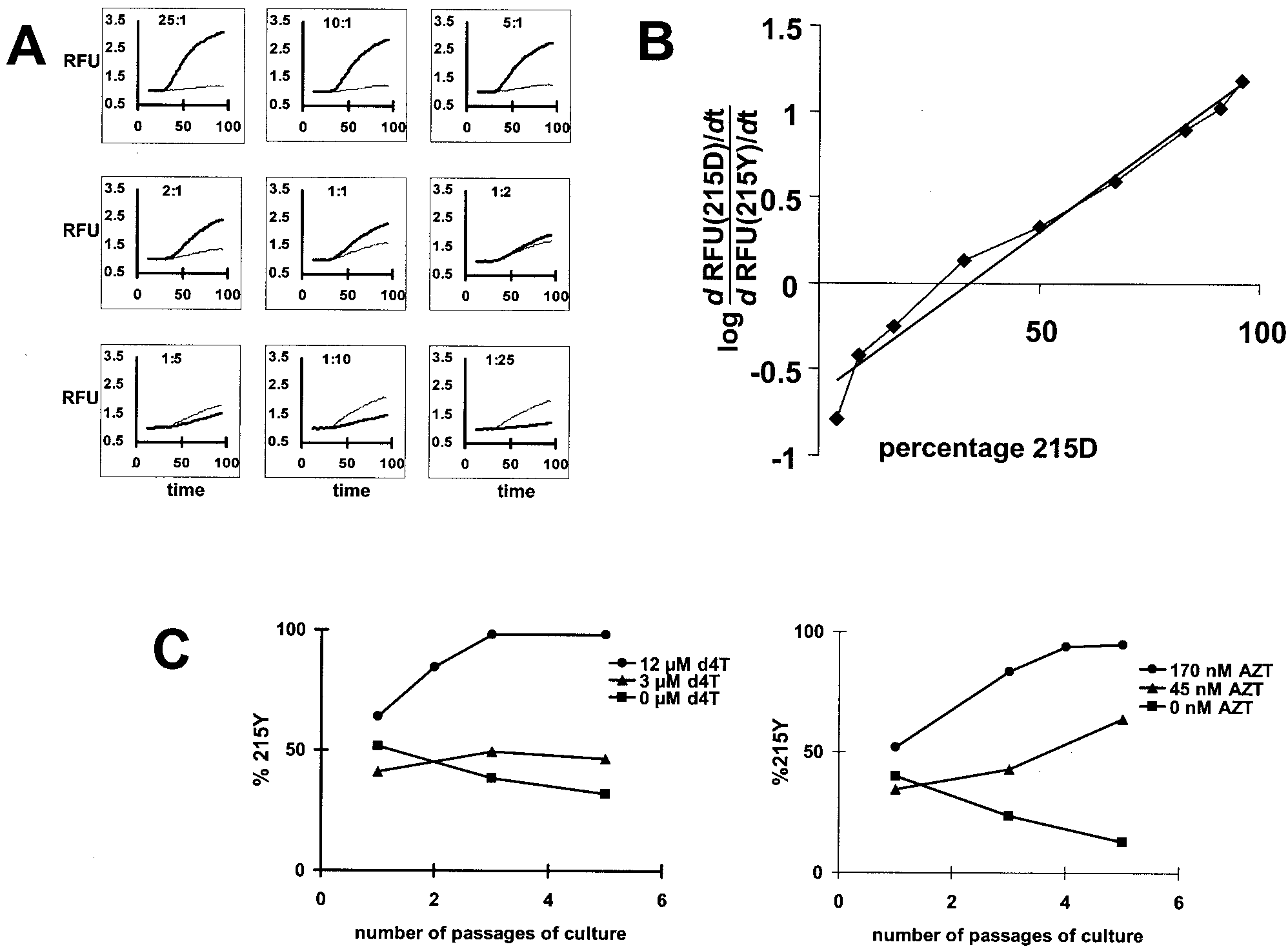 FIG. 4. In vitro competitions of 215Y and 215D viruses in the absence and presence of antiviral drugs as analyzed by using real-time NASBA.
FIG. 4. In vitro competitions of 215Y and 215D viruses in the absence and presence of antiviral drugs as analyzed by using real-time NASBA.