Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Pii: s0031-9422(01)00191-
Stefan Martensa, Gert Forkmanna, Ulrich Maternb,*, Richard Lukacˇinb
aTechnische Universita¨t Mu¨nchen, Wissenschaftszentrum fu¨r Erna¨hrung, Landnutzung und Umwelt, Department fu¨r Pflanzenwissenschaften,
Lehrstuhl fu¨r Zierpflanzenbau, Am Hochanger 4, D-85350 Freising, Germany
bInstitut fu¨r Pharmazeutische Biologie, Philipps-Universita¨t Marburg, Deutschhausstrasse 17 A, D-35037 Marburg, Germany
Received 19 March 2001; received in revised form 6 April 2001
A cDNA encoding flavone synthase I was amplified by RT-PCR from leaflets of Petroselinum crispum cv. Italian Giant seedlings
and functionally expressed in yeast cells. The identity of the recombinant, 2-oxoglutarate-dependent enzyme was verified in assaysconverting (2S)-naringenin to apigenin. # 2001 Elsevier Science Ltd. All rights reserved.
Keywords: Petroselinum crispum; Apiaceae; Flavonoid biosynthesis; Flavone synthase I cloning; 2-Oxoglutarate-dependent dioxygenase; Hetero-logous expression
During the last decade considerable progress has been
cephalosporin biosynthesis (Baldwin and Abraham,
achieved towards elucidating the mode of action and
1988), as well as in mammalian tissues (Lindstedt et al.,
molecular architecture of 2-oxoglutarate-dependent diox-
1977; Kivirikko et al., 1989; Stenflo et al., 1989). Fur-
ygenases. These enzymes catalyze diverse reactions, such
thermore, these enzymes catalyze numerous reactions in
as the hydroxylation, desaturation, epoxidation or cycli-
plants, e.g. in the formation of hydroxyproline-rich gly-
zation of substrates, and the activities depend on ferrous
coproteins (Tanaka et al., 1980), of gibberellins (Hed-
iron and molecular oxygen which is reduced during cata-
den and Graebe, 1982) and the secondary metabolites
lysis by two electrons provided by decarboxylation of the
scopolamine (Hashimoto and Yamada, 1986), vindoline
cosubstrate (Prescott, 1993; DeCarolis and DeLuca, 1994;
(DeCarolis et al., 1990) or of various flavonoids (Fork-
Prescott and John, 1996). Although 2-oxoglutarate is the
mann et al., 1980; Britsch et al., 1981; Lukacˇin and
common cosubstrate, some closely related dioxygenases
Britsch, 1997; Lukacˇin et al., 2000 a,b,c).
mobilize the electrons from decarboxylation of the sub-
Five 2-oxoglutarate-dependent dioxygenases have been
strate itself, e.g. isopenicillin N-synthase (Baldwin and
identified so far from flavonoid biosynthesis, which
Abraham, 1988) and 4-hydroxyphenylpyruvate dioxy-
include the widely distributed anthocyanidin synthase
genase (Bradley et al., 1986; Ru¨etschi et al., 1992), or by
(Menssen et al., 1990), flavanone 3b-hydroxylase (Fork-
the oxidation of ascorbate as in ethylene biosynthesis
mann et al., 1980; Britsch et al., 1981; Lukacˇin and
(Zhang et al., 1995; Lay et al., 1996). These latter
Britsch, 1997; Lukacˇin et al., 2000 a,b,c) and flavonol
enzymes may nevertheless classify with the 2-oxogluta-
synthase (Britsch et al., 1981; Holton et al., 1993).
rate-dependent dioxygenases stricto sensu in one cate-
Another dioxygenase, catalyzing the 6-hydroxylation of
gory of intermolecular dioxygenases. Intermolecular
partially methylated flavonols, was reported recently
dioxygenases fulfill a variety of pivotal functions in pri-
from Chrysosplenium americanum (Anzelotti and Ibra-
mary and secondary metabolism in bacteria (Omura et
him, 2000), while flavone synthase I, FNS I, appears to
al., 1984; Salowe et al., 1990) and fungi, including the
be confined to species of the Apiaceae (Britsch, 1990).
cyclization and ring expansion reactions in penicillin/
FNS I had been characterized in 1981 as a solubleenzyme from parsley, in contrast to the microsomal fla-vone synthase II, FNS II, from other plants (Kochs andGrisebach, 1987; Martens and Forkmann, 1998), and
* Corresponding author. Tel.: +49-6421-282-2461; fax: +49-6421-
was partially purified through six-steps of fractionation
E-mail address: matern@mailer.uni-marburg.de (U. Matern).
from irradiated cell cultures (Britsch, 1990). This
0031-9422/01/$ - see front matter # 2001 Elsevier Science Ltd. All rights reserved. P I I : S 0 0 3 1 - 9 4 2 2 ( 0 1 ) 0 0 1 9 1 - 1
S. Martens et al. / Phytochemistry 58 (2001) 43–46
enzyme was then employed in kinetic studies aiming at
1992), and the similarity of prolyl 4-hydroxylase with
the reaction mechanism, which revealed that synthetic
lysyl hydroxylase from chicken (Myllyla¨ et al., 1991) or
2-hydroxynaringenin did not compete with flavanone
of fungal isopenicillin N-synthase with desacetoxy-
substrates. Accordingly, the 2,3-desaturation of flava-
cephalosporin C synthase ranged only at approx. 20%
nones by FNS I was postulated to proceed by direct
(Britsch et al., 1993). Nevertheless, superimposing the
abstraction of the vicinal hydrogen atoms (Britsch,
structural models of the penicillin and cephalosporin
1990), which would assign FNS I to a distinct desatur-
synthases revealed an almost identical architecture for
ase subgroup among the 2-oxoglutarate-dependent
these two enzymes (Lloyd et al., 1999), and comparison
of the CD spectra of Petunia flavanone 3b-hydroxylase
exluding the successive hydroxylation and dehydrata-
and isopenicillin N-synthase suggested the same pattern
tion, was proposed for the desaturation of alkanes to
of helical, non-helical and b-sheet motifs for the Petunia
olefines suggesting a reaction via radical intermediates
dioxygenase (Lukacˇin et al., 2000b). Flavanone 3b-
(Mansuy, 1998). The exact course of FNS I catalysis
hydroxylase and FNS I both use 2-oxoglutarate as the
requires further experimental support, but appears to
cosubstrate and depend on the same flavanone sub-
proceed analogous to that of the cytochrome P450-
strates (Fig. 1). Accordingly, a thorough examination of
dependent FNS II expressed in many plants except for
the sequential and spatial differences of these two
the Apiaceae. The first full size FNS II cDNAs were
enzymes, together with in vitro mutagenesis studies,
recently cloned from Gerbera hybrida (Martens and
might be rather helpful to pinpoint the putative sub-
Forkmann, 1999), Antirrhinum majus and Torenia
strate binding sites and to explain the formation of fla-
hybrida (Akashi et al., 1999) by differential display PCR
and expressed in yeast cells. As anticipated for a P450-
Based on alignments of fourteen intermolecular diox-
dependent monoxygenase, this FNS II converted label-
ygenase polypeptides from public data bases two con-
led flavanones to the corresponding flavones apparently
served sequence motifs were chosen, and, similar to the
without any intermediate (Martens and Forkmann,
previous cloning of flavonol synthase (Fig. 1) from Pet-
unia hybrida (Holton et al., 1993), degenerate oligonu-
The common mode of oxygen activation by inter-
cleotide primers were designed for the cloning of FNS I.
molecular dioxygenases, particularly among the 2-oxo-
In combination with oligo(dT), the primers were
glutarate-dependent enzymes, seems to predict a high
employed for RT-PCR amplification of cDNAs from
degree of homology at the DNA and polypeptide levels.
total RNA that had been extracted from young leaflets
However, only 30% similarity was observed in the
at four stages of development of flavone-producing
polypeptide sequences of, for example, flavanone 3b-
Petroselinum crispum cv. Italian Giant plants (Martens
hydroxylase from Petunia hybrida and hyoscyamine 6b-
and Forkmann, 1999). A whole set of intermolecular
hydroxylase from Hyoscyamus niger (Britsch et al.,
dioxygenase cDNAs was amplified, and the full-size
Fig. 1. Reaction catalyzed by flavone synthase I (FNS I), converting (2S)-naringenin to apigenin, in comparison to the activities of flavanone 3b-hydroxylase (FHT) and flavonol synthase (FLS), which sequentially convert (2S)-naringenin to dihydrokaempferol and kaempferol.
S. Martens et al. / Phytochemistry 58 (2001) 43–46
unpublished). Moreover, the yeast strain transfectedwith the pYES2 vector hosting the FNS I cDNA in theinverse orientation did not express flavone synthaseactivity (Fig. 2 B). Recombinant FNS I lacked flavonolsynthase activity (Fig. 1), and the sequences of these twoenzymes differ considerably. Thus, soluble FNS I pre-vailing in the Apiaceae was cloned for the first time andhas become available in quantity for mechanistic studiesas well as for the convenient preparative synthesis ofradiolabeled flavones which enable further biosyntheticand biotechnological studies. The evolutionary contextfor the expression of the soluble synthase exclusively inthe Apiaceae remains to be established. In addition, therecombinant enzyme may be of value for the productionof flavone-nutraceuticals due to their antioxidant andanticancer potentials (Harborne and Williams, 2000).
We are indebted to Dr. L. Britsch, Dr. R. Zimmer-
mann and Dr. H. Mu¨ller (Merck KGaA, Darmstadt)for the ESI–MS analysis of apigenin as the product ofthe recombinant parsley FNS I.
Akashi, T., Fukuchi-Mizutani, M., Aoki, T., Ueyama, Y., Yonekura-
Fig. 2. Catalytic activity of parsley FNS I expressed in yeast cells.
Sakakibara, K., Tanaka, Y., Kusumi, T., Ayabe, S., 1999. Mole-
Crude extracts from yeast cells expressing the FNS I from the cDNA
cular cloning and biochemical characterization of novel cytochrome
inserted in the pYES2 expression vector (A) or harbouring the FNS I
P450, flavone synthase II, that catalyzes direct conversion of flava-
cDNA in the inverse orientation (B) were incubated with (2S)-[4a,6-
nones to flavones. Plant Cell Physiology 40, 1182–1186.
8-14C]naringenin (NAR) as described for the native plant enzyme
Anzelotti, D., Ibrahim, R., 2000. Novel flavonol 2-oxoglutarate-depen-
(Britsch, 1990). Subsequently, the incubations were extracted with
dent dioxygenase: affinity purification, characterization, and kinetic
ethyl acetate, the extracts were separated by thin-layer chromatograpy
properties. Archives of Biochemistry and Biophysics 382, 161–172.
on cellulose in 30% aqueous acetic acid (v/v), and the radioactivity
Baldwin, J.E., Abraham, E., 1988. The biosynthesis of penicillins and
was spotted by a phosphorimager (Martens and Forkmann, 1998).
cephalosporins. Natural Product Reports 5, 129–145.
The product was identified by cochromatography with authentic api-
Bradley, F.C., Lindstett, S., Lipscomb, J.D., Que Jr., L., Roe, A.L.,
Rundgren, M., 1986. 4-Hydroxyphenylpyruvate is an iron-tyr-osinate protein. Journal of Biological Chemistry 261, 11693–11696.
Britsch, L, Heller, W., Grisebach, H., 1981. Conversion of flavanone to
clones were generated by the 50-RACE technique. In
flavone, dihydroflavonol and flavonol with an enzyme system from cell
addition to flavanone 3b-hydroxylase, flavonol syn-
cultures of parsley. Zeitschrift fu¨r Naturforschung 36c, 742–750.
thase, 1-aminocyclopropane-1-carboxylate oxidase and
Britsch, L., 1990. Purification and characterization of flavone synthase
two not yet fully characterized 2-oxoglutarate-depen-
I, a 2-oxoglutarate-dependent desaturase. Archives of Biochemistryand Biophysics 276, 348–354.
dent dioxygenases, FNS I was also recognized among
Britsch, L., Ruhnau-Brich, B., Forkmann, G., 1992. Molecular clon-
the cDNA clones. The FNS I cDNA was unequivocally
ing, sequence analysis, and in vitro expression of flavanone 3b-
identified by expression in yeast strain INVSc1, using
hydroxylase from Petunia hybrida. Journal of Biological Chemistry
the expression vector pYES2 (Invitrogen, Groningen,
The Netherlands), and the efficient conversion of nar-
Britsch, L., Dedio, J., Saedler, H., Forkmann, G., 1993. Molecular
characterization of flavanone 3b-hydroxylase: consensus sequence,
ingenin to apigenin by the recombinant enzyme (Fig. 2
comparison with related enzymes and the role of conserved histi-
A). The identity of the reaction product was confirmed
dines. European Journal of Biochemistry 217, 745–754.
by direct comparison of the retention time on reversed
DeCarolis, E., Chan, F., Balsevich, J., DeLuca, V., 1990. Isolation
phase HPLC in water/acetonitrile 7:3 and the ESI–MS
and characterization of a 2-oxoglutarate-dependent dioxygenase
spectrum with the mobility and fragmentation pattern
involved in the second-to-last step in vindoline biosynthesis. PlantPhysiology 94, 1323–1329.
of authentic apigenin (data not shown). The FNS I
DeCarolis, E., DeLuca, V., 1994. 2-Oxoglutarate-dependent dioxy-
polypeptide was also recognized in Western blots by a
genase and related enzymes: biochemical characterization. Phyto-
FNS I polyclonal rabbit antiserum (Lukacˇin et al.,
S. Martens et al. / Phytochemistry 58 (2001) 43–46
Forkmann, G., Heller, W., Grisebach, H., 1980. Anthocyanin bio-
Mansuy, D., 1998. The great diversity of reactions catalyzed by cyto-
synthesis in flowers of Matthiola incana. Flavanone 3- and flavonoid
chromes P450. Comparative Biochemistry and Physiology Part C
30-hydroxylases. Zeitschrift fu¨r Naturforschung 35c, 691–695.
Harborne, J.B., Williams, C.A., 2000. Advances in flavonoid research
Martens, S., Forkmann, G., 1998. Genetic control of flavone synthase
since 1992. Phytochemistry 55, 481–504.
II activity in flowers of Gerbera hybrids. Phytochemistry 49, 1953–
Hashimoto, T., Yamada, Y., 1986. Hyoscyamine 6b-hydroxylase, a 2-
oxoglutarate-dependent dioxygenase, in alkaloid producing root
Martens, S., Forkmann, G., 1999. Cloning and expression of flavone
cultures. Plant Physiology 81, 619–625.
synthase II from Gerbera hybrids. Plant Journal 20, 611–618.
Hedden, P., Graebe, J.E., 1982. Cofactor requirements for the soluble
Menssen, A., Ho¨hmann, S., Martin, W., Schnable, P.S., Peterson,
oxidases in the metabolism of the C20-gibberellins. Journal of Plant
P.A., Saedler, H., Gierl, A., 1990. The En/Spm transposable ele-
ment of Zea mays contains splice sites at the termini generating a
Holton, T.A., Brugliera, F., Tanaka, Y., 1993. Cloning and expression of
novel intron from a dSpm element in the A2 gene. EMBO Journal 9,
flavonol synthase from Petunia hybrida. Plant Journal 4, 1003–1010.
Kivirikko, K.I., Myllyla¨, R., Pihlajaniemi, T., 1989. Protein hydro-
Myllyla¨, R., Pihlajaniemi, T., Pajunen, L., Turpeenniemi-Hujanen, T.,
xylation: prolyl 4-hydroxylase, an enzyme with four cosubstrates
Kivirikko, K.I., 1991. Molecular cloning of chick lysyl hydroxylase.
and a multifunctional subunit. FASEB Journal 3, 1609–1617.
Journal of Biological Chemistry 266, 2805–2810.
Kochs, G., Grisebach, H., 1987. Induction and characterization of a
Omura, S., Tomoda, H., Yamamoto, S., Tsukui, M., Tanaka, H.,
NADPH-dependent flavone synthase from cell cultures of soybean.
1984. Studies on two dioxygenases involved in the synthesis of
Zeitschrift fu¨r Naturforschung 42c, 343–348.
tylosin in Streptomyces fradiae. Biochimica et Biophysica Acta 802,
Lay, V.J., Prescott, A.G., Thomas, P.G., John, P., 1996. Heterologous
expression and site directed mutagenesis of the 1-aminocyclopro-
Prescott, A.G., 1993. A dilemma of dioxygenases (or where biochem-
pane-1-carboxylate oxidase from kiwi fruit. European Journal of
istry and molecular biochemistry fail to meet). Journal of Experi-
Lindstedt, G., Lindstedt, S., Nordin, I., 1977. Purification and prop-
Prescott, A.G., John, P., 1996. Dioxygenases: molecular structure and
erties of g-butyrobetaine hydroxylase from Pseudomonas sp AK 1.
role in plant metabolism. Annual Reviews of Plant Physiology and
Plant Molecular Biology 47, 245–271.
Lloyd, M.D., Lee, H.-J., Harlos, K., Zhang, Z.-H., Baldwin, J.E.,
Ru¨etschi, U., Odelho¨g, B., Lindstedt, S., Barros-So¨derling, J., Persson,
Schofield, C.J., Charnock, J.M., Garner, C.D., Hara, T., Terwisscha
B., Jo´rnvall, H., 1992. Characterization of 4-hydroxyphenyl-
van Scheltinga, A.C., Valega˚rd, K., Viklund, J.A.C., Hajdu, J.,
pyruvate dioxygenase: primary structure of the Pseudomonas
Andersson, I., Danielsson, A˚., Bhikhabhai, R., 1999. Studies on the
enzyme. European Journal of Biochemistry 205, 459–466.
active site of deacetoxycephalosporin C synthase. Journal of Mole-
Salowe, S.P., Marsh, E.N., Townsend, C.A., 1990. Purification and
characterization of clavaminate synthase from Streptomyces clavu-
Lukacˇin, R., Britsch, L., 1997. Identification of strictly conserved his-
ligerus an unusual oxidative enzyme in natural product biosynthesis.
tidine and arginine residues as part of the active site in Petunia
hybrida flavanone 3b-hydroxylase. European Journal of Biochem-
Stenflo, J., Holme, E., Lindstedt, S., Chandramouli, N., Tsai Huang,
L.H., Tam, J.P., Merrifield, R.B., 1989. Hydroxylation of aspartic
Lukacˇin, R., Gro¨ning, I., Schiltz, E., Britsch, L., Matern, U., 2000a.
acid in domains homologous to the epidermal growth factor pre-
Purification of recombinant flavanone 3b-hydroxylase from Petunia
cursor is catalyzed by a 2-oxoglutarate-dependent dioxygenase.
hybrida and assignment of the primary site of proteolytic degrada-
Proceedings of the National Academy of Sciences USA 86, 444–447.
tion. Archive of Biochemistry and Biophysics 375, 364–370.
Tanaka, M., Shibata, H., Uchida, T., 1980. A new prolyl hydroxylase
Lukacˇin, R., Gro¨ning, I., Pieper, U., Matern, U., 2000b. Site-directed
acting on poly-L-proline from suspension cultered cells of Vinca
mutagenesis of the active site serine 290 in flavanone 3b-hydroxylase
rosea. Biochimica et Biophysica Acta 616, 188–198.
from Petunia hybrida. European Journal of Biochemistry 267, 853–860.
Zhang, Z., Schofield, C.J., Baldwin, J.E., Thomas, P., John, P., 1995.
Lukacˇin, R., Urbanke, C., Gro¨ning, I., Matern, U., 2000c. The
Expression, purification and characterization of 1-aminocyclopro-
monomeric polypeptide comprises the functional flavanone 3b-
pane-1-carboxylate oxidase from tomato in Escherichia coli. Bio-
hydroxylase from Petunia hybrida. FEBS Letters 467, 353–358.
ANA/NJ Newsletter Vol X, No. 2 April 2006 Meeting at East Brunswick Public Library, February 19, 2006 This was a meeting for sharing experiences, conducted by Kathie Belonger, who also provided the delicious refreshments. There were 15 people in attendance. In the course of an informal and wide-ranging discussion, we learned about (1) coping with an AN back in the 1970s, and dea
SUMMARY OF CHANGES TO QOF 2014/15 - ENGLAND KEY No change Indicator Business Exception 14/15 QOF NICE ID Indicator wording Threshold Threshold timeframe timeframe timeframe (months) (months) (months) CLINICAL Atrial Fibrilation (AF) The contractor establishes and maintains a register of patients with atrial The percentage of patients w

 Stefan Martensa, Gert Forkmanna, Ulrich Maternb,*, Richard Lukacˇinb
aTechnische Universita¨t Mu¨nchen, Wissenschaftszentrum fu¨r Erna¨hrung, Landnutzung und Umwelt, Department fu¨r Pflanzenwissenschaften,
Lehrstuhl fu¨r Zierpflanzenbau, Am Hochanger 4, D-85350 Freising, Germany
bInstitut fu¨r Pharmazeutische Biologie, Philipps-Universita¨t Marburg, Deutschhausstrasse 17 A, D-35037 Marburg, Germany
Received 19 March 2001; received in revised form 6 April 2001
A cDNA encoding flavone synthase I was amplified by RT-PCR from leaflets of Petroselinum crispum cv. Italian Giant seedlings
and functionally expressed in yeast cells. The identity of the recombinant, 2-oxoglutarate-dependent enzyme was verified in assaysconverting (2S)-naringenin to apigenin. # 2001 Elsevier Science Ltd. All rights reserved.
Stefan Martensa, Gert Forkmanna, Ulrich Maternb,*, Richard Lukacˇinb
aTechnische Universita¨t Mu¨nchen, Wissenschaftszentrum fu¨r Erna¨hrung, Landnutzung und Umwelt, Department fu¨r Pflanzenwissenschaften,
Lehrstuhl fu¨r Zierpflanzenbau, Am Hochanger 4, D-85350 Freising, Germany
bInstitut fu¨r Pharmazeutische Biologie, Philipps-Universita¨t Marburg, Deutschhausstrasse 17 A, D-35037 Marburg, Germany
Received 19 March 2001; received in revised form 6 April 2001
A cDNA encoding flavone synthase I was amplified by RT-PCR from leaflets of Petroselinum crispum cv. Italian Giant seedlings
and functionally expressed in yeast cells. The identity of the recombinant, 2-oxoglutarate-dependent enzyme was verified in assaysconverting (2S)-naringenin to apigenin. # 2001 Elsevier Science Ltd. All rights reserved.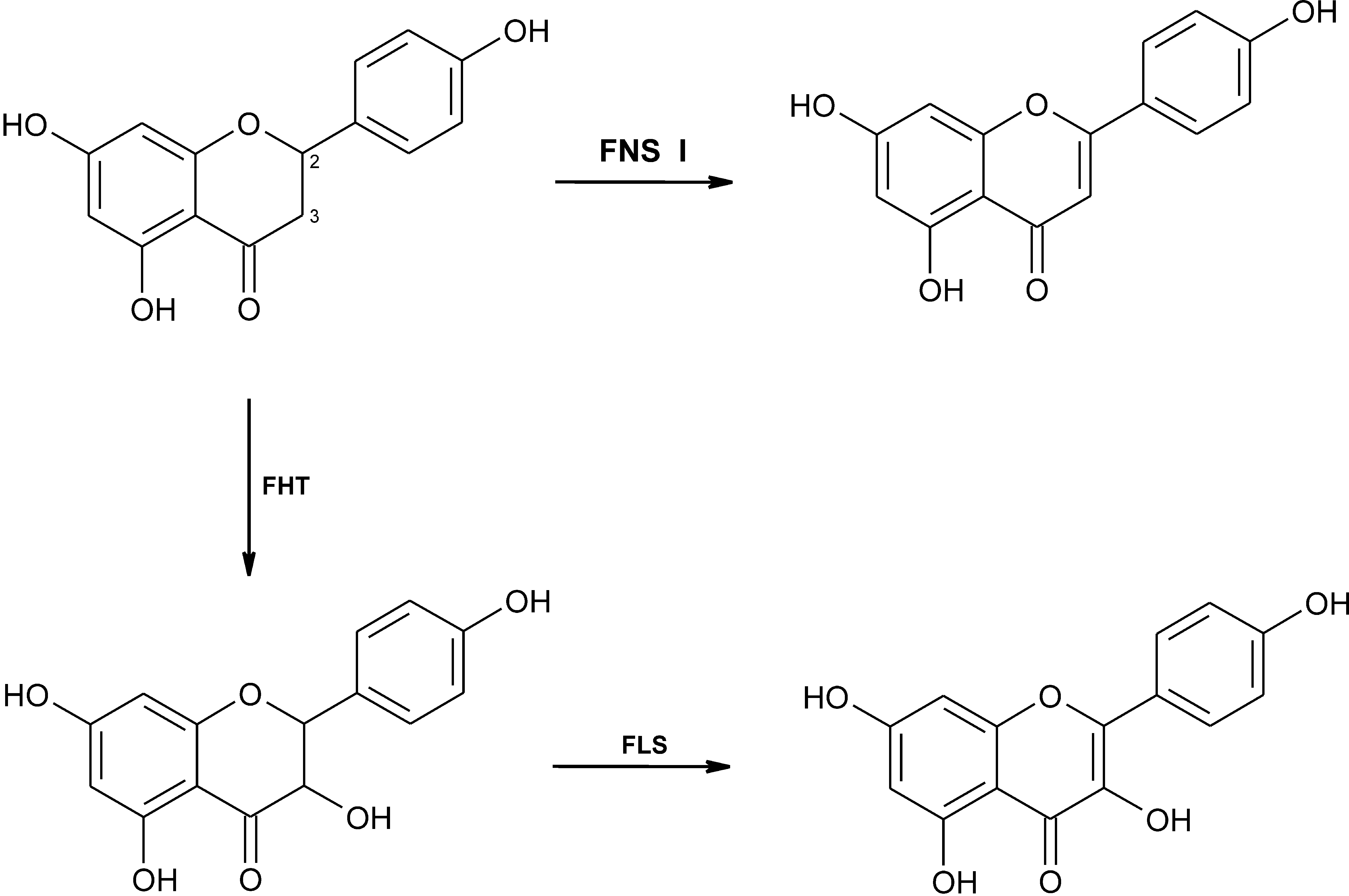 S. Martens et al. / Phytochemistry 58 (2001) 43–46
enzyme was then employed in kinetic studies aiming at
1992), and the similarity of prolyl 4-hydroxylase with
the reaction mechanism, which revealed that synthetic
lysyl hydroxylase from chicken (Myllyla¨ et al., 1991) or
2-hydroxynaringenin did not compete with flavanone
of fungal isopenicillin N-synthase with desacetoxy-
substrates. Accordingly, the 2,3-desaturation of flava-
cephalosporin C synthase ranged only at approx. 20%
nones by FNS I was postulated to proceed by direct
(Britsch et al., 1993). Nevertheless, superimposing the
abstraction of the vicinal hydrogen atoms (Britsch,
structural models of the penicillin and cephalosporin
1990), which would assign FNS I to a distinct desatur-
synthases revealed an almost identical architecture for
ase subgroup among the 2-oxoglutarate-dependent
these two enzymes (Lloyd et al., 1999), and comparison
of the CD spectra of Petunia flavanone 3b-hydroxylase
exluding the successive hydroxylation and dehydrata-
and isopenicillin N-synthase suggested the same pattern
tion, was proposed for the desaturation of alkanes to
of helical, non-helical and b-sheet motifs for the Petunia
olefines suggesting a reaction via radical intermediates
dioxygenase (Lukacˇin et al., 2000b). Flavanone 3b-
(Mansuy, 1998). The exact course of FNS I catalysis
hydroxylase and FNS I both use 2-oxoglutarate as the
requires further experimental support, but appears to
cosubstrate and depend on the same flavanone sub-
proceed analogous to that of the cytochrome P450-
strates (Fig. 1). Accordingly, a thorough examination of
dependent FNS II expressed in many plants except for
the sequential and spatial differences of these two
the Apiaceae. The first full size FNS II cDNAs were
enzymes, together with in vitro mutagenesis studies,
recently cloned from Gerbera hybrida (Martens and
might be rather helpful to pinpoint the putative sub-
Forkmann, 1999), Antirrhinum majus and Torenia
strate binding sites and to explain the formation of fla-
hybrida (Akashi et al., 1999) by differential display PCR
and expressed in yeast cells. As anticipated for a P450-
Based on alignments of fourteen intermolecular diox-
dependent monoxygenase, this FNS II converted label-
ygenase polypeptides from public data bases two con-
led flavanones to the corresponding flavones apparently
served sequence motifs were chosen, and, similar to the
without any intermediate (Martens and Forkmann,
previous cloning of flavonol synthase (Fig. 1) from Pet-
unia hybrida (Holton et al., 1993), degenerate oligonu-
The common mode of oxygen activation by inter-
cleotide primers were designed for the cloning of FNS I.
S. Martens et al. / Phytochemistry 58 (2001) 43–46
enzyme was then employed in kinetic studies aiming at
1992), and the similarity of prolyl 4-hydroxylase with
the reaction mechanism, which revealed that synthetic
lysyl hydroxylase from chicken (Myllyla¨ et al., 1991) or
2-hydroxynaringenin did not compete with flavanone
of fungal isopenicillin N-synthase with desacetoxy-
substrates. Accordingly, the 2,3-desaturation of flava-
cephalosporin C synthase ranged only at approx. 20%
nones by FNS I was postulated to proceed by direct
(Britsch et al., 1993). Nevertheless, superimposing the
abstraction of the vicinal hydrogen atoms (Britsch,
structural models of the penicillin and cephalosporin
1990), which would assign FNS I to a distinct desatur-
synthases revealed an almost identical architecture for
ase subgroup among the 2-oxoglutarate-dependent
these two enzymes (Lloyd et al., 1999), and comparison
of the CD spectra of Petunia flavanone 3b-hydroxylase
exluding the successive hydroxylation and dehydrata-
and isopenicillin N-synthase suggested the same pattern
tion, was proposed for the desaturation of alkanes to
of helical, non-helical and b-sheet motifs for the Petunia
olefines suggesting a reaction via radical intermediates
dioxygenase (Lukacˇin et al., 2000b). Flavanone 3b-
(Mansuy, 1998). The exact course of FNS I catalysis
hydroxylase and FNS I both use 2-oxoglutarate as the
requires further experimental support, but appears to
cosubstrate and depend on the same flavanone sub-
proceed analogous to that of the cytochrome P450-
strates (Fig. 1). Accordingly, a thorough examination of
dependent FNS II expressed in many plants except for
the sequential and spatial differences of these two
the Apiaceae. The first full size FNS II cDNAs were
enzymes, together with in vitro mutagenesis studies,
recently cloned from Gerbera hybrida (Martens and
might be rather helpful to pinpoint the putative sub-
Forkmann, 1999), Antirrhinum majus and Torenia
strate binding sites and to explain the formation of fla-
hybrida (Akashi et al., 1999) by differential display PCR
and expressed in yeast cells. As anticipated for a P450-
Based on alignments of fourteen intermolecular diox-
dependent monoxygenase, this FNS II converted label-
ygenase polypeptides from public data bases two con-
led flavanones to the corresponding flavones apparently
served sequence motifs were chosen, and, similar to the
without any intermediate (Martens and Forkmann,
previous cloning of flavonol synthase (Fig. 1) from Pet-
unia hybrida (Holton et al., 1993), degenerate oligonu-
The common mode of oxygen activation by inter-
cleotide primers were designed for the cloning of FNS I.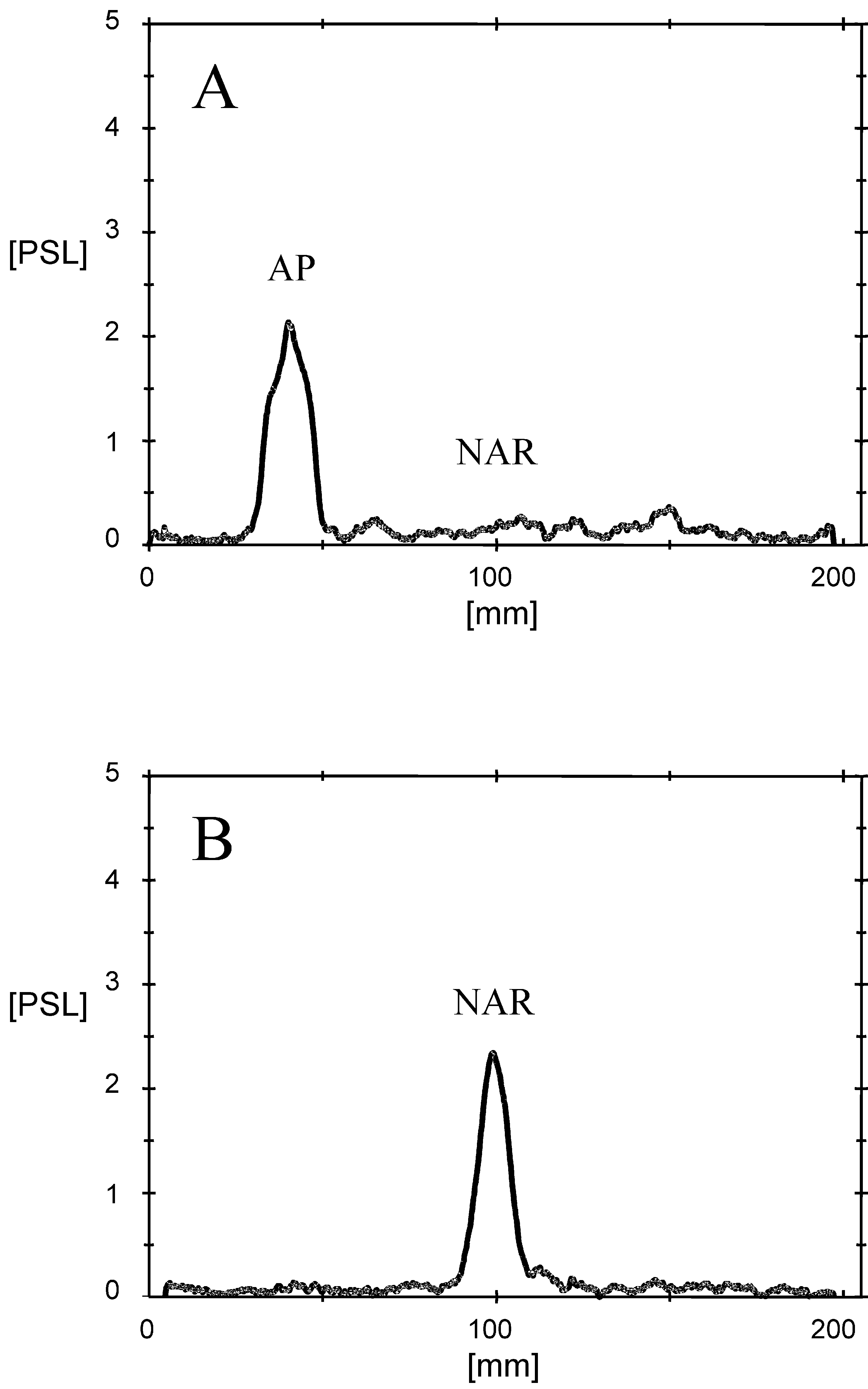 S. Martens et al. / Phytochemistry 58 (2001) 43–46
unpublished). Moreover, the yeast strain transfectedwith the pYES2 vector hosting the FNS I cDNA in theinverse orientation did not express flavone synthaseactivity (Fig. 2 B). Recombinant FNS I lacked flavonolsynthase activity (Fig. 1), and the sequences of these twoenzymes differ considerably. Thus, soluble FNS I pre-vailing in the Apiaceae was cloned for the first time andhas become available in quantity for mechanistic studiesas well as for the convenient preparative synthesis ofradiolabeled flavones which enable further biosyntheticand biotechnological studies. The evolutionary contextfor the expression of the soluble synthase exclusively inthe Apiaceae remains to be established. In addition, therecombinant enzyme may be of value for the productionof flavone-nutraceuticals due to their antioxidant andanticancer potentials (Harborne and Williams, 2000).
S. Martens et al. / Phytochemistry 58 (2001) 43–46
unpublished). Moreover, the yeast strain transfectedwith the pYES2 vector hosting the FNS I cDNA in theinverse orientation did not express flavone synthaseactivity (Fig. 2 B). Recombinant FNS I lacked flavonolsynthase activity (Fig. 1), and the sequences of these twoenzymes differ considerably. Thus, soluble FNS I pre-vailing in the Apiaceae was cloned for the first time andhas become available in quantity for mechanistic studiesas well as for the convenient preparative synthesis ofradiolabeled flavones which enable further biosyntheticand biotechnological studies. The evolutionary contextfor the expression of the soluble synthase exclusively inthe Apiaceae remains to be established. In addition, therecombinant enzyme may be of value for the productionof flavone-nutraceuticals due to their antioxidant andanticancer potentials (Harborne and Williams, 2000).