Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Hartnell.cc.ca.us
2000 Nature America Inc. • http://neurosci.nature.com articles Early sleep triggers memory for early visual discrimination skills
Steffen Gais, Werner Plihal, Ullrich Wagner and Jan Born
Clinical Neuroendocrinology, Medical University of Lübeck, Ratzeburger Allee 160/Hs 23a, 23538, Lübeck, GermanyCorrespondence should be addressed to J.B. (born@kfg.mu-luebeck.de)Improvement after practicing visual texture discrimination does not occur until several hours after practice has ended. We show that this improvement strongly depends on sleep. To specify the process responsible for sleep-related improvement, we compared the effects of ‘early’ and ‘late’ sleep, dominated respectively by slow-wave and rapid eye movement (REM) sleep. Discrimination skills significantly improved over early sleep, improved even more over a whole night’s sleep, but did not improve after late sleep alone. These findings suggest that procedural memory formation is prompted by slow-wave sleep-related processes. Late REM sleep may promote memory formation at a second stage, only after periods of early sleep have occurred. osci.nature
The consolidation of memories, a concept introduced a century
retrieval testing, thus making it difficult to make straightforward
ago1, is particularly supported by processes occurring during
conclusions about the consolidation process22,23. To avoid these
sleep. The earliest studies on this topic2,3 and later animal and
problems, we dissociated functions of REM sleep and SWS for
• http://neur
human studies4–6 provide considerable evidence that sleep helps
visual procedural memory by splitting the night into two halves9.
to consolidate memories. Spatiotemporal patterns of neuronal
In humans, the first half of sleep is normally dominated by peri-
activity are replayed in the rat hippocampus during periods of
ods of SWS, and there is little REM sleep. During late sleep, this
slow-wave sleep (SWS) following learning. This replay has been
pattern reverses. If REM sleep is essential for consolidation of
linked to consolidation of declarative types of (spatial) memory
discrimination skills, consolidation should be strengthened from
occurring during this sleep stage7,8. Consistent with these find-
a retention period encompassing predominantly REM sleep,
ings, human memory for word pairs and spatial locations bene-
rather than SWS, and vice versa. Accordingly, learning of texture
fits significantly more from early sleep dominated by extended
discrimination skills was compared after nocturnal retention
SWS than from late sleep where REM sleep prevails9–11. Where-
periods of either early or late sleep. In control conditions, sub-
2000 Nature America Inc.
as these observations pertain to a declarative type of memory
jects remained awake throughout corresponding retention peri-
that relies mainly on the integrity of the hippocampus and adja-
ods. Improvement in texture discrimination was measured by
cent temporal lobe structures, attention has only recently been
comparing the minimum presentation time (stimulus to mask
focused on non-declarative, procedural types of memory10–13.
onset asynchrony, SOA) necessary to discriminate orientation of
Procedural knowledge refers to implicit (or pre-attentive) learn-
a target feature, before and after the retention period.
ing of ‘habits’ or ‘how to’ memories by practicing sensory andmotor skills, respectively14. Unlike the declarative memory sys-
tem, procedural memory does not necessarily involve hip-
Sleep data confirmed that SWS dominated early sleep and REM
pocampal functions but, depending on the type of task, relies on
sleep dominated late sleep (Table 1). During initial learning
various neocortical and subcortical structures15,16.
before sleep, texture discrimination performance did not differ
The performance improvement for a basic texture discrimi-
between the early and late conditions (122 ± 6.7 ms versus
nation task takes place in assemblies of neurons active at a very
120 ± 6.3 ms, p > 0.7). Subjects were retested after sleep, and
early pre-attentive stage of visual processing17. Substantial
their discrimination skills improved only after early sleep. Dur-
improvement in perceptual performance of this task occurs eight
ing late sleep and during both early and late wake intervals, dis-
or more hours after it has ended, rather than during or immedi-
crimination performance even decreased, that is, threshold SOA
ately after practice; this finding indicates a slow, latent process of
increased (Fig. 1a). This pattern was statistically confirmed by
learning18. Sleeping during the retention interval seems to be
overall ANOVA with two within-group factors—threshold SOA
particularly important for improvement19. Selective disruption
before versus after the retention interval, and early versus late
of REM sleep blocks overnight enhancement of the perceptual
retention interval—and one between-group factor, sleep versus
skill, whereas SWS disruption has no effect on enhancement12.
wake (F1,25 = 7.23, p < 0.02 for before/after × sleep/wake inter-
These results led to the conclusion that the consolidation of tex-
action; F1,25 = 9.74, p < 0.01 for main effect of before/after). Sep-
ture discrimination, that is, enhancing the neural pathways
arate analysis of the early and late conditions revealed this
involved in this task and thereby improving task performance, is
interaction to be significant for the early condition (F1,13 = 9.73,
a process strongly dependent on REM sleep. p < 0.01). Performance improvement, as measured by the
However, the REM sleep deprivation protocol has been cri-
decrease in threshold SOA, was significant after early sleep
tiqued20,21 because it induces distinct emotional and cognitive
(t7 = 2.33, p < 0.05), whereas wake controls showed an oppos-
disturbances that interfere with task performance at the time of
ing tendency toward increasing thresholds across the early reten-
nature neuroscience • volume 3 no 12 • december 2000 2000 Nature America Inc. • http://neurosci.nature.com articles
tion interval (t6 = –2.22, p < 0.10). For
Table 1. Sleep stages by experimental condition. Early sleep Late sleep t
interval in both the sleep and wakegroups, we found a significant main
p < 0.02). Separate analysis of sleep
nificant before/after × early/late inter-
Time (in minutes, and in percent of total sleep time; mean ± s.e.m.) spent in each sleep stage during early and
action (F1,13 = 8.20, p < 0.02) for
late sleep. Right column, results for pairwise statistical analysis by t-test. *p < 0.001.
sleeping subjects and a significantmain effect of before/after(F1,12 = 10.39, p < 0.01) for the wake control group, thus con-
mance on the well-learned task at 2200 hours, 0300 hours and
firming a selective improvement in discrimination skills after the
0800 hours was compared to performance on a task with a new
early retention sleep, and a decrease in performance especially
target location, which differed at each test. The subjects’ perfor-
mance was comparable for all three times of testing, for familiar
In a supplementary study, we examined the improvement in
stimuli (82 ± 14 ms, 79 ± 10 ms and 86 ± 17 ms respectively) and
texture discrimination after a 12-hour retention interval, which
for novel stimuli (122 ± 26 ms, 120 ± 23 ms and 134 ± 23 ms),
either was at night and contained an 8-hour period of sleep, or
excluding essential effects of circadian rhythm on discrimination
was during the day and did not contain a period of sleep. As
skills (p > 0.50 for all comparisons). Performance on familiar
expected from previous studies18, the threshold SOA in this
stimuli was better than performance on novel stimuli (p < 0.001
experiment decreased from 130 ± 14.3 ms before sleep to
105 ± 12.1 ms after sleep (t5 = 6.07, p < 0.001) in those subjects
osci.nature
tested over a night retention interval. However, subjects’ perfor-
DISCUSSION
mance did not improve during the day retention interval of equal
Data from the main study showed that texture discrimination skills
length (141 ± 10.9 ms before versus 143 ± 13.4 ms after,
during the night improved only if the retention interval contained
t5 = –0.337, p > 0.7). In addition, the improvement in discrimi-
SWS-dominated early sleep; during late sleep alone, threshold SOA
• http://neur
nation skill during the full period of nocturnal sleep was com-
even increased. Texture discrimination skills also deteriorated when
pared with that observed in the main experiment during a period
subjects were kept awake during the retention phase (main effect
of early sleep alone. Whereas task performance before sleep was
across wake conditions). This pattern cannot be explained by cir-
comparable for both of these conditions (130 ± 14.3 ms versus
cadian rhythms because discrimination skills improved when sub-
122 ± 6.7 ms, p > 0.5), the improvement during the entire night
jects slept during the early retention interval but did not improve
was about three times greater than the improvement during early
when they were kept awake during this time. In addition, circadi-
sleep alone (t12 = –3.23, p < 0.01; Fig. 1b).
an rhythms did not influence performance on a well-learned or
A second supplementary study focused on the possible influ-
novel discrimination task. Because the wake control group was
ences of circadian rhythm on discrimination performance, and
deprived of sleep, their fatigue at retrieval testing may have
2000 Nature America Inc.
tested subjects on a well-learned task. As expected, discrimina-
impaired texture discrimination performance. However, if the
tion task training led to an asymptotic reduction in threshold
improvement in texture discrimination was only due to sleep before
SOA (first session, 115 ± 7 ms; tenth session, 85 ± 6 ms). Perfor-
retrieval testing, improvement should also have occurred after late
Fig. 1. Improvements in visual discrimination skill. (a) Difference in threshold SOA between learning and retrieval testing, after retention intervals during the early or late half of the night (mean ± s.e.m.). Subjects either slept for three hours during the retention interval, or were kept awake throughout the time interval. Asterisk, p < 0.05, for difference between conditions. Threshold SOA decreased only across the early sleep retention condition. †, p < 0.05, for a comparison with learning performance before the retention interval. (b) Difference in threshold SOA between learning and retrieval testing after the early sleep retention interval (white bar), the 12-hour retention intervals of a whole night’s sleep (black bar), and the awake daytime period (gray bar). Larger improvement in visual discrimination was observed after a whole night’s sleep, than after a period of early sleep alone. **p < 0.01 for dif- ference between conditions.
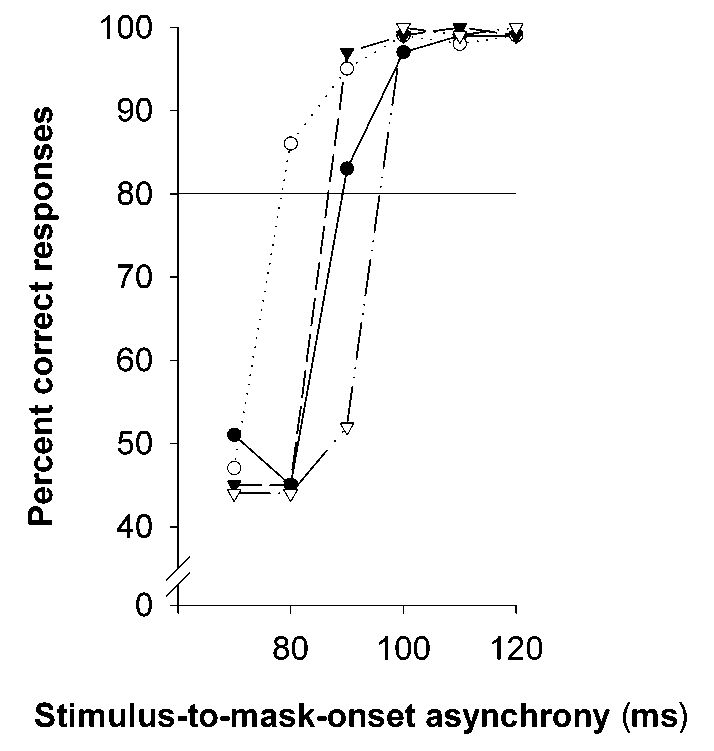
††p < 0.01 for a comparison with learning performance before the retention interval. (c) Individual performance curves for two subjects tested on the early sleep condition (circles) and late sleep condi- tion (triangles). Filled symbols, performance at initial learning before sleep; open symbols, performance at retrieval test- ing after sleep. Horizontal line, 80% correct responses. Threshold SOA is the point where performance curves cross the horizontal line. nature neuroscience • volume 3 no 12 • december 2000 2000 Nature America Inc. • http://neurosci.nature.com articles Fig. 2. Experimental protocol. Schedule (a) for the ‘early’ and ‘late’ sleep condition and (b) for the ‘early’ and ‘late’ wake control condition. Blank fields, times when subjects were awake; gray fields, intervals of sleep; black fields, time of texture discrimination task.
sleep. Moreover, initial task performance was very similar before
Comparable amounts of time were spent in REM sleep during
early and late retention intervals, which argues against any con-
the REM sleep deprivation condition of that study (19 ± 6 min)
siderable influence of fatigue or circadian variation.
and the early sleep condition of our study (24.3 ± 4.1 min). The
osci.nature
The finding that performance improved only after early sleep
amounts of time spent in SWS during the SWS deprivation com-
suggests that processes related to this period of sleep facilitate con-
ponent of the previous study (30 ± 12 min) and during the late
solidation of these procedural skills. The predominance of SWS
sleep condition of our study (31.6 ± 4.1 min) were also compa-
and associated cortical changes in excitability24 and transmitter
rable. However, the REM sleep deprivation protocol of the pre-
• http://neur
turnover25 may be an essential prerequisite for this facilitation.
vious study differed from our approach, in that this stage was
Besides SWS dominance, early sleep is also characterized by var-
disrupted after the REM sleep process was initiated. REM sleep
ious neurohormonal changes, such as an inhibition of pituitary-
deprivation leads to substantial fragmentation of sleep architec-
adrenal release, which may be involved in memory consolidation.
ture. Resulting from the frequent arousals during REM sleep,
Although studies in rodents indicate memory enhancement for
emotional as well as attentive disturbances can be observed6,22,23.
emotionally aversive tasks after administration of glucocorticoids
These disturbances particularly affect retrieval testing perfor-
(particularly into the basolateral amygdala26), human studies with
mance after sleep20,21. Considering our finding of improved tex-
systemic administration of glucocorticoids during early noctur-
ture discrimination after early sleep, it is difficult to determine
nal sleep consistently show an impairment of declarative memo-
why SWS deprivation did not disturb consolidation12. Howev-
2000 Nature America Inc.
ry function during this period27,28.
er, in the previous experiments, despite repetitive arousals, sub-
The present findings contrast with results from a previous
jects spent an average of 30 minutes in SWS, in which the
experiment, in which selective deprivation of REM sleep pre-
consolidation process may have been initiated29.
vented an improvement in texture discrimination skill across
A supplementary experiment addressed two further issues.
nocturnal sleep, and deprivation of SWS sleep had no effect12.
First, it indicated that sleep is necessary after practice to stimulate
Fig. 3. Task Displays. Example of a stimulus (a) and a mask (b). All line positions varied slightly from trial to trial and the ‘T’ or ‘L’ in the center of the screen was rotated randomly. The target texture (three diagonal lines), upper left quadrant of the stimulus display (a). nature neuroscience • volume 3 no 12 • december 2000 2000 Nature America Inc. • http://neurosci.nature.com articles
any consistent improvement in visual discrimination. When sub-
synaptic long-term potentiation and activity-dependent synap-
jects were tested during the daytime wake phase, some subjects
togenesis known to depend on modulatory influences from cer-
improved, whereas others deteriorated in performance. The fail-
tain neurotransmitters and neurohormonal inputs35–39.
ure of wake retention intervals to stimulate significant improve-
Experimental variation of transmitter and neurohormonal lev-
ment in discrimination skills confirms previous data18,19. Second,
els during selected sleep periods is a possible approach to deter-
when improvement after a full night of sleep was compared with
mining the synaptic mechanisms involved in the different steps of
improvement observed in the main experiment following early
sleep alone, a striking interdependence of early and late sleep wasrevealed. The improvement across the entire sleep period was
about three times greater than the improvement after early sleep
Subjects were healthy non-smokers (19–35 years old), with normal or
alone (Fig. 1a). This difference was not only attributable to the
corrected to normal vision. They slept seven to nine hours per night, and
longer retention interval across the full night, because a reten-
had no major disruptions of the sleep–wake cycle during the six weeks
tion interval of equal length without sleep had no effect on task
before experimentation. They were not allowed to ingest caffeine or alco-hol, or sleep during the day before experimental nights. Before the exper-
performance. Rather, the better performance after a full night of
iment, subjects were accustomed to sleeping under laboratory conditions.
sleep suggests a two-step consolidation process.
The experiments were approved by the Ethics Committee of the Med-
Thus, although our data do not suggest that REM sleep is of
primary importance for the learning of visual discrimination
In the main experiment, each of 15 subjects participated in two exper-
skills, REM sleep may add to consolidating memories once the
imental nights, which were about one week apart. On these nights, sub-
effects of early sleep have been manifested. A previous correla-
jects learned a texture discrimination task with the retention interval
tional study19 used the same discrimination task as our study,
between learning and retrieval testing encompassing either the early or
and showed that the improvement in texture discrimination skills
late half of the night. Subjects were randomly assigned to either a sleep
is correlated with the amount of time spent in SWS in the begin-
group (n = 8), which had a 3-hour period of sleep during the retention
ning of the night, and the time spent in REM sleep toward the
interval, or a control group, which remained awake during the retention
osci.nature
end of the night. The present data extends this finding, and pro-
All sleep and wake periods could vary to a limited degree, to individually
vides the first experimental evidence for a two-step process of
adapt them to a subject’s normal sleep–wake rhythm (Fig. 2), but eight
memory formation during sleep, in which the second, REM
hours between learning and retrieval testing were required, because oth-
sleep-related step is only effective after memory processes have
erwise, no improvement could be expected for the task18. Sleep time was
• http://neur
been initiated in a first SWS-related step. Our experimental obser-
measured from sleep onset. Subjects were woken during the first stage-two
vations show that REM-rich late sleep alone is ineffective for
sleep occurring after three hours of sleep. During the time the subjects
memory consolidation, and that visual discrimination skills after
were awake, they were not allowed to visually or physically strain them-
an eight-hour sleep period (containing normal amounts of both
selves. Most of the time, they played board games or listened to music.
SWS and REM sleep) are, on average, more than three times more
Two supplementary experiments were done to control for effects of cir-
cadian rhythm on retention and discrimination performance, respectively.
improved than after a period of early sleep alone. The view of a
In the first experiment, 12-hour retention intervals between learning and
two-step memory consolidation process would also integrate
retrieval testing encompassed either nighttime (2100 hours–0900 hours;
findings that arousals during REM sleep deteriorate consolida-
n = 6) or daytime (0900 hours–2100 hours; n = 6). The nighttime inter-
tion of texture discrimination skills after early sleep and SWS
val included a complete 8-hour sleep interval beginning at 2300 hours. 2000 Nature America Inc.
have initiated consolidation12. Once memory traces have gained
In the second supplementary experiment, seven subjects were tested on
some strength, neocortical stimulation during REM sleep could
a well-learned task. They reported to the laboratory for 10 consecutive
lead to a reactivation of previously encoded materials, sharpen-
days, practicing the discrimination task with the target displayed in one
ing the traces30,31. Whether the small amount of REM sleep dur-
location. Afterward, subjects spent one experimental night in the labora-
ing early sleep (as well as the small amount of SWS in late sleep)
tory, being tested for discrimination performance at 2200 hours,
have a particular function in this kind of sequential consolida-
0300 hours and 0800 hours. They slept two times for three hours duringthese nights (2300–0200 hours and 0400–0700 hours). At each test, per-
tion process remains to be determined.
formance on the well-learned task was compared to performance on a
Based on its local (retinotopic) nature, texture discrimination
novel discrimination task, with the target displayed in a new location.
is considered to take place early during visual processing in the
The visual discrimination task was designed as described17. It was car-
primary visual cortex and closely associated areas17,32. The learn-
ried out in a silent and dark environment. Stimuli were presented on a
ing of texture discrimination occurs at a pre-attentive level, and
Macintosh PowerPC computer with a 17-inch monitor (75 Hz). Subjects
hence represents a fundamental type of procedural memory. In
were asked to react by pressing keys on a keyboard. Each session con-
this regard, the selective improvement in discrimination skills
sisted of 1250 trials, each composed of three sequential displays. First, a
after a period of early sleep diverges from previous studies in
cross was displayed in the center of the screen. Subjects were told to leave
which a greater enhancement of procedural memory was
their eyes fixed at this point throughout the trial. After they pressed akey, there was a blank screen interval of 250–300 ms. Second, the target
observed after periods of late rather than early retention
display (Fig. 3a) was shown for 10 ms, followed by another blank screen
sleep10,11,13. One of those studies used a mirror-tracing task; the
interval. Third, the mask was presented for 100 ms (Fig. 3b). Exposure
other used a word-stem priming task. Those tasks seem to be
to the mask overrode the remains of the target display on the retina. Thus,
more complex than the texture discrimination task. The influ-
discrimination difficulty could be systematically increased by reducing
ence of REM sleep may become increasingly important with more
the stimulus to mask onset asynchrony (SOA).
complex tasks13,33,34, although the neurophysiological meaning of
The target displays were 16° of visual angle in size and contained a field
‘task complexity’ in this context remains to be specified.
of 19 × 19 horizontal bars with a randomly rotated ‘T’ or ‘L’ shaped figure
The neurophysiological mechanisms underlying sleep-asso-
in the center. The target (three horizontally or vertically aligned diagonal
ciated facilitation of procedural memories are unclear. Improve-
bars) was located in the peripheral visual field at a distance of 3°–5° of
ment in texture discrimination skills may be a result of
visual angle from the center. After each trial, subjects had to report by keypress the letter in the center of the display and the target orientation, that
use-dependent changes in cell connectivity within V1 and close-
is, the alignment of the three diagonal lines. Discrimination of the cen-
ly connected areas17,32. These changes might involve processes of
nature neuroscience • volume 3 no 12 • december 2000 2000 Nature America Inc. • http://neurosci.nature.com articles
tral letter assured correct eye fixation, and only trials with correct letter
13. Aubrey, J., Smith, C., Tweed, S. & Nader, R. Cognitive and motor procedural
discrimination were used for analysis. There was no time limit for giving
tasks are dissociated in REM and stage two sleep. Sleep Res. Online 2, 220
the answer, and subjects were told to take breaks when needed. SOA was
14. Squire, L. R. & Zola, S. M. Structure and function of declarative and
continuously reduced after a block of trials. Steps were 460, 360, 260 and
nondeclarative memory systems. Proc. Natl. Acad. Sci.USA 93, 13515–13522
220 ms with 50 trials per step. Then SOA was reduced from 180 to 60 ms
in steps of 20 ms, with 150 trials at each step. Thus, a total of 1250 trials
15. Pascual-Leone, A., Grafman, J. & Hallett, M. Modulation of cortical motor
were done for each period of learning and retrieval testing. Before the
output maps during development of implicit and explicit knowledge. Science 263, 1287–1289 (1994).
experiment, subjects practiced the task in the presence of the experimenter
16. Pascual-Leone, A. et al. Procedural learning in Parkinson’s disease and
with 50 trials at a SOA of 460 ms. Performance was measured as the per-
cerebellar degeneration. Ann. Neurol. 34, 594–602 (1993).
cent correct responses at a given SOA. Threshold SOA was estimated by
17. Karni, A. & Sagi, D. Where practice makes perfect in texture discrimination:
interpolating the point where the recognition rate was 80%. Thus,
evidence for primary visual cortex plasticity. Proc. Natl. Acad. Sci.USA 88,
improvement in texture discrimination between learning and retrieval
18. Karni, A. & Sagi, D. The time course of learning a visual skill. Nature 365,
testing was equivalent to a reduction in threshold SOA. Sessions with a
threshold SOA of more than 220 ms were excluded from analysis, because
19. Stickgold, R., Whidbee, D., Schirmer, B., Patel, V. & Hobson, J. A. Visual
they were clearly recognizable as outliers.
discrimination task improvement: A multi-step process occurring during
The target was shown in the same quadrant of the visual field during
sleep. J. Cogn. Neurosci. 12, 246–254 (2000).
20. Vertes, P. & Eastman, K. The case against memory consolidation in REM
learning and subsequent retrieval testing after a retention period, but
sleep. Behav. Brain Sci. (in press).
different quadrants were used on the two experimental conditions of a
21. Born, J. & Gais, S. REM sleep deprivation: the wrong paradigm leading to
subject. Because improvement in this task is retinally local, that is, it
wrong conclusions. Behav. Brain Sci. (in press).
occurs only within the area of the visual field where the target is repeat-
22. Horne, J. A. & McGrath, M. J. The consolidation hypothesis for REM sleep
edly presented17, improvement measured in two different quadrants of
function: stress and other confounding factors—a review. Biol. Psychol. 18, 165–184 (1984).
the visual field can be assumed to be independent. This was confirmed by
23. Oniani, T. N. Does paradoxical sleep deprivation disturb memory trace
consolidation? Physiol. Behav. 33, 687–692 (1984).
24. Marshall, L., Molle, M., Fehm, H. L. & Born, J. Scalp recorded direct current
brain potentials during human sleep. Eur. J. Neurosci. 10, 1167–1178 (1998). ACKNOWLEDGEMENTS
25. McCormick, D. A. Neurotransmitter actions in the thalamus and cerebral
osci.nature
cortex. J. Clin. Neurophysiol. 9, 212–223 (1992). We thank A. Karni and R. Stickgold for providing the software for the texture
26. Roozendaal, B. Glucocorticoids and the regulation of memory consolidation. discrimination task, and for comments on the manuscript. This research wasPsychoneuroendocrinology 25, 213–238 (2000). supported by a grant from the Deutsche Forschungsgemeinschaft (DFG) to J.B.
27. Plihal, W., Pietrowsky, R. & Born, J. Dexamethasone blocks sleep induced
improvement of declarative memory. Psychoneuroendocrinology 24, 313–331 (1999). • http://neur RECEIVED 21 AUGUST; ACCEPTED 20 SEPTEMBER 2000
28. Plihal, W. & Born, J. Memory consolidation in human sleep depends on
inhibition of glucocorticoid release. Neuroreport 10, 2741–2747 (1999).
29. Born, J., Muth, S. & Fehm, H. L. The significance of sleep onset and slow wave
1. Müller, G. E. & Pilzecker, A. Experimentelle beiträge zur lehre vom
sleep for nocturnal release of growth hormone (GH) and cortisol.
gedächtnis. Z. Psychol. 1, 1–300 (1900). Psychoneuroendocrinology 13, 233–243 (1988).
2. Heine, R. Uber wiedererkennen und ruckwirkende hemmung. Z. Psychol. 68,
30. Giuditta, A. et al. The sequential hypothesis of the function of sleep. Behav.Brain Res. 69, 157–166 (1995).
3. Jenkins, J. G. & Dallenbach, K. M. Obliviscence during sleep and waking. Am.
31. Maquet, P. et al. Reactivation during rapid eye movement (REM) sleep of
J. Psychol. 35, 605–612 (1924).
cerebral areas involved in the execution of a serial reaction time (SRT) task.
4. Hennevin, E., Hars, B., Maho, C. & Bloch, V. Processing of learned
Sleep Res. Online 2, 694 (1999).
information in paradoxical sleep: relevance for memory. Behav. Brain Res. 69,
32. Schoups, A. A. & Orban, G. A. Interocular transfer in perceptual learning of a
pop-out discrimination task. Proc. Natl. Acad. Sci. USA 93, 7358–7362
5. Smith, C. Sleep states and memory processes. Behav. Brain Res. 69, 137–145 2000 Nature America Inc.
33. Empson, J. A. & Clarke, P. R. Rapid eye movements and remembering.
6. Cipolli, C. Sleep, dreams and memory: an overview. J. Sleep Res. 4, 2–9 Nature 227, 287–288 (1970).
34. Smith, C. Sleep states, memory processes and synaptic plasticity. Behav. Brain
7. Wilson, M. A. & McNaughton, B. L. Reactivation of hippocampal ensemble
Res. 78, 49–56 (1996).
memories during sleep. Science 265, 676–679 (1994).
35. Kirkwood, A., Rioult, M. C. & Bear, M. F. Experience-dependent
8. Buzsaki, G. Memory consolidation during sleep: a neurophysiological
modification of synaptic plasticity in visual cortex. Nature 381, 526–528
perspective. J. Sleep Res. 7, 17–23 (1998).
9. Yaroush, R., Sullivan, M. J. & Ekstrand, B. R. Effect of sleep on memory. II.
36. Kirkwood, A., Rozas, C., Kirkwood, J., Perez, F. & Bear, M. F. Modulation of
Differential effect of the first and second half of the night. J. Exp. Psychol. 88,
long-term synaptic depression in visual cortex by acetylcholine and
norepinephrine. J. Neurosci. 19, 1599–1609 (1999).
10. Plihal, W. & Born, J. Effects of early and late nocturnal sleep on declarative
37. Edagawa, Y., Saito, H. & Abe, K. Serotonin inhibits the induction of long-
and procedural memory. J. Cogn. Neurosci. 9, 534–547 (1997).
term potentiation in rat primary visual cortex. Prog. Neuropsychopharmacol.
11. Plihal, W. & Born, J. Effects of early and late nocturnal sleep on priming and
Biol. Psychiatry 22, 983–997 (1998).
spatial memory. Psychophysiology 36, 571–582 (1999).
38. McEwen, B. S. Neurosteroids. (eds. Baulieu, E. E., Robel, P. & Schumacher,
12. Karni, A., Tanne, D., Rubenstein, B. S., Askenasy, J. J. & Sagi, D. Dependence
M.) 233–254 (Humana, Totowa, New Jersey, 1999).
on REM sleep of overnight improvement of a perceptual skill. Science 265,
39. Cahill, L. & McGaugh, J. L. Modulation of memory storage. Curr. Opin.Neurobiol. 6, 237–242 (1996). nature neuroscience • volume 3 no 12 • december 2000
MILVERTON C. P. SCHOOL First Aid Policy We have three trained First Aiders in school:- All our trained first aiders are experienced in dealing with ailments and accidents and must be consulted on First Aid matters. Medical Care Parents/carers of children who need to take medicines during the school day are asked to bring the medicine to the medical room at the beginning of the sc
Gemeindezeitung Winter 2007 FIEBER Die durchschnittliche Körpertemperatur des Menschen beträgt 37,2°Celsius und ist am Nachmittag um ca. 17h am höchsten. Man spricht von beginnendem Fieber (subfebril) unter 38°C, mäßigem Fieber bis 39°C, und hohem Fieber über 39°C bis hin zur höchsten Körpertemperatur von 43° C. Fieber ist eine Abwehrreaktion des Körpers und ist daher der ent

 2000 Nature America Inc. • http://neurosci.nature.com
2000 Nature America Inc. • http://neurosci.nature.com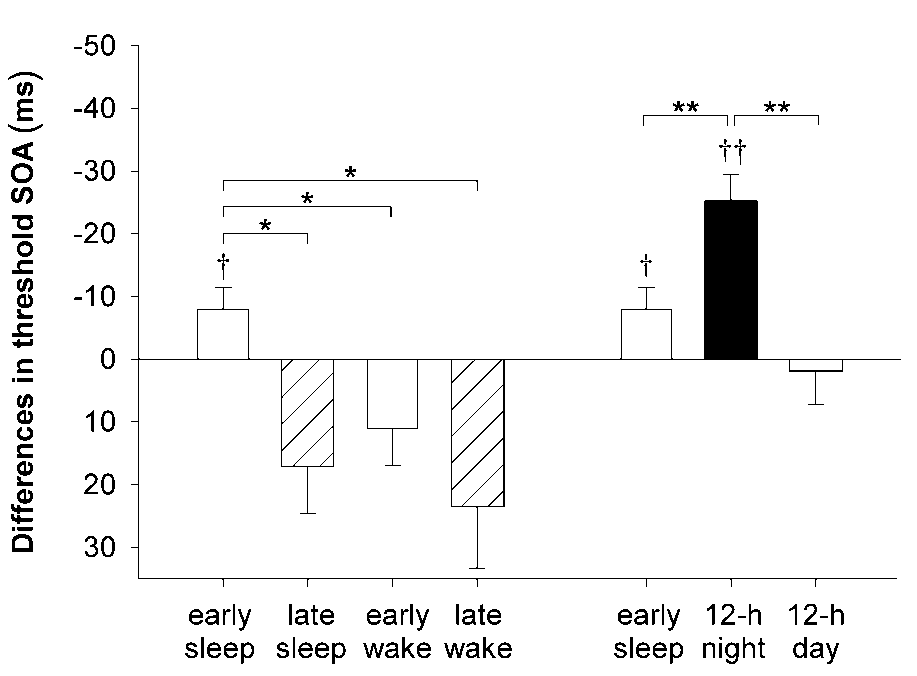

 2000 Nature America Inc. • http://neurosci.nature.com
2000 Nature America Inc. • http://neurosci.nature.com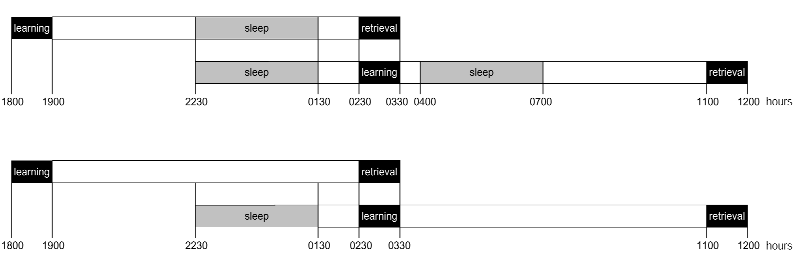


 2000 Nature America Inc. • http://neurosci.nature.com
2000 Nature America Inc. • http://neurosci.nature.com
 2000 Nature America Inc. • http://neurosci.nature.com
2000 Nature America Inc. • http://neurosci.nature.com
 2000 Nature America Inc. • http://neurosci.nature.com
2000 Nature America Inc. • http://neurosci.nature.com