Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Doi:10.1016/j.clinph.2004.04.015
Clinical Neurophysiology 115 (2004) 2157–2163
Fluoxetine facilitates use-dependent excitability
Burkhard Pleger*, Peter Schwenkreis, Christian Gru¨nberg, Jean-Pierre Malin, Martin Tegenthoff
Department of Neurology, Ruhr-University Bochum, BG-Kliniken Bergmannsheil, Buerkle-de-la-Camp-Platz 1, D-44789 Bochum, Germany
Objectives: In poststroke patients, fluoxetine, a selective serotonin-reuptake inhibitor, as an adjunct to physical therapy provided a better
functional recovery from motor deficits. The aim of this study was to investigate the effect of a single dose of 20 mg fluoxetine on motorlearning and associated cortical changes in healthy right-handed subjects in order to get deeper insight into its facilitating influence on humanmotor cortex.
Methods: Subjects performed a motor task consisting of a simultaneous co-contraction of the abductor pollicis brevis (APB) and the
deltoid muscle with and without fluoxetine in a placebo-controlled double-blinded crossover study design. Immediately before and aftermotor learning motor output maps of the APB muscle were assessed in order to get insight into plastic changes of the musclerepresentation.
Results: We found a significantly improved motor performance under both conditions without having substantial differences between
placebo and fluoxetine. After the completion of the motor task there was a medial shift of the APB muscle motor output map. Only afterthe administration of fluoxetine the sum of MEP amplitudes (SOA) increased and the motor output map enlarged.
Conclusions: These findings provide evidence for a use-dependent facilitating effect of fluoxetine on cortical excitability but not on
Significance: Our findings are not in line with previous experiments in poststroke patients. However, long-term treatment with
fluoxetine may additionally improve motor function by upregulating serotonergic receptors. Further studies investigating the influence oflong-term treatment on cortical excitability and psychophysics may therefore provide deeper insight into a possible therapeuticalefficiency of fluoxetine in poststroke patients.
q 2004 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved.
Keywords: Transcranial magnetic stimulation; Primary motor cortex; Plasticity; Fluoxetine; Serotonin
(). Among otherantidepressants, it inhibits 5-HT reuptake selectively, with
The application of the endogenous transmitter 5-hydro-
no anticholinergic activity and limited side-effects (
xytrytamine (5-HT, serotonin) into cell cultures causes
long-term facilitation of sensorimotor synapses due in part
an adjunct to physical therapy provided a better functional
to growth of new presynaptic varicosities (
recovery from motor deficits (). In first
). In several animal experiments, 5-HT showed the
functional magnetic resonance imaging experiments,
capacity to modulate purposeful motor responses
Pariente et al. found a fluoxetine-dependent hyperactivation
in the ipsilesional primary motor cortex giving rise to an
enhanced hemodynamic response of the perilesional tissue
inhibitor fluoxetine accumulates in the brain relative to the
plasma and promotes an amplified serotonin concentration
In the present study we investigated the effect of a
single dose of 20 mg fluoxetine on psychophysical and
* Corresponding author. Tel.: þ 49-234-3026094; fax: þ 49-234-
cortical changes in healthy right-handed subjects in order
to get deeper insight into its facilitating influence on
E-mail address: burkhard.v.pleger@ruhr-uni-bochum.de (B. Pleger).
1388-2457/$30.00 q 2004 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.clinph.2004.04.015
B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
human motor system. In previous studies synchronised
movement of two limb muscles led to rapid plasticchanges of the motor output map
TMS was applied using a figure-of-eight-shaped coil
(outside diameter 8.7 cm, peak magnetic field strength
the present study subjects performed a so-called
2.2 T, peak electric field strength 660 V/m) that was
connected with a Magstim 200 HP device (monopolar
contraction of the abductor pollicis brevis (APB) and
waveform/The Magstim Company). The TMS measurement
was performed immediately before and after the co-con-
traction task. The subjects were seated comfortably in a
double-blinded crossover study design. In all subjects
chair. Motor evoked potentials (MEP) were recorded on an
motor output map of the APB muscle was assessed by
EMG device (sampling rate: 5000 Hz/Neuropack 8, Nihon
transcranial magnetic stimulation (TMS) of the primary
Kohden) with surface electrodes from the APB muscle of
the dominant hand. The band pass filter was set from 20 Hzto 2 kHz, the gain from 0.1 to 1 mV/D. The sufficientmuscle relaxation during the examination was continuously
monitored by EMG (gain 0.1 mV/D). The TMS coil washeld tangentially to the head in an anterior – posterior
direction, with the grip pointing backwards. Motor threshold(MT) was defined as the minimum intensity which produced
We examined ten healthy right-handed subjects (five men
five motor evoked potentials . 50 mV out of ten trials. It was
and four women, aged 25 – 29 years, 27.3 ^ 3.3 years,
determined over that scalp position were TMS previously
mean ^ standard error). They all gave their written
elicited the highest amplitude. Stimulation intensity was set
informed consent, and the protocol of the study was
to 110% of the motor threshold. Starting at the scalp
approved by the local ethical committee.
position of the highest amplitudes, the motor cortex was
The study was performed in a placebo-controlled
examined in dorsal, rostral, lateral and medial direction in
double-blinded crossover design. The subjects had to
steps of 1 cm until no further MEP could be elicited. For a
participate in two sessions. Therefore, the supervisor
systematic examination of the primary motor cortex we used
(M.T.) randomly divided them into two groups, each
a tight fitting cap with a coordinate system on it (1 £ 1 cm
consisting of five subjects. The first group started after the
width). The coordinate system was arranged relative to Cz,
administration of a single dose of 20 mg fluoxetine, whereas
which was identified as the intersection of the interaural line
the subjects of the second group first received placebo.
ðy-axis), and the connection between nasion and inion
In line with previous co-contraction studies (
ðx-axis). Eight stimuli were applied to each position, and the
responses (peak-to-peak amplitudes) were averaged. These
the groups were reversed after an interval of two
averaged amplitudes were considered for further statistical
weeks in order to avoid a carry over effect. The investigators
analysis. Averaged amplitudes smaller than 10 mV were
did not know if the subjects had taken placebo or fluoxetine
rejected. Afterwards, we calculated the sum of all
prior to each session. The randomization code was kept by
MEP-amplitudes of the motor output map (SOA), and its
the supervisor and broken at the end of the study after the
amplitude-weighted centre of gravity (COG). The centre of
individual data analysis had been completed.
gravity (COG) is a single x – y coordinate derived from the
Under both conditions, subjects had to perform a motor
distribution of MEP amplitudes within the motor output
task 6 h after drug intake to guarantee peak plasma
area. It was calculated according to the following formula:
concentration after fluoxetine administration (
the number of positions from which MEPs could be
consisted of a synchronised movement of the abductor
elicited was used as a marker for the area size of the
pollicis brevis (APB) muscle and the deltoid muscle (for a
detailed description see ). The subjectswere instructed to make brisk and short movements of
both muscles as synchronously as possible. Over 1 h,three co-contractions per minute had to be performed.
To evaluate differences of the TMS parameters assessed
During each single co-contraction we measured the
under placebo and fluoxetine we used an ANOVA for
latency difference between the onsets of both muscle
repeated measurements (pre vs. post training) with the
contractions using EMG-monitoring from surface electro-
within-subject factor ‘condition’ (placebo vs. fluoxetine).
des. We used the differences between the muscle onsets as
For post-hoc analysis we then used the student’s paired
a marker of motor performance. After each co-contraction
t-test to compare the TMS parameters obtained before and
the subjects were informed of the results and encouraged
after the task within each session. We also checked
the reproducibility of the neurophysiological data
B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
by comparing the maps obtained prior to motor learning of
12.41 ^ 1 ms; P , 0:001). Using post-hoc t-test, we found
significantly reduced time differences between the first
To evaluate the effect of repetitive co-contraction on
and the successive sessions (1 – 10 vs. 11 – 20: P ¼ 0:003;
motor performance, the mean latency differences between
1 – 10 vs. 21 – 30: P ¼ 0:0003; 1–10 vs.31 –40: P ¼ 0:001;
the onsets of both muscles for the intervals 0 – 10, 10 – 20,
20 – 30, 30 – 40, 40 – 50 and 50 – 60 min were calculated in
P ¼ 0:0004). Using the ANOVA for repeated measure-
each subject and for each session. We used an ANOVA
ments and the within-subject factor ‘time £ condition’,
for repeated measurements and post-hoc t-test analysis
we found no differences between placebo and fluoxetine
(Bonferroni-corrected for multiple comparisons) to express
ðFð5;80Þ ¼ 0:59; P ¼ 0:7; ‘time’: Fð5;80Þ ¼ 39:59; P , 0:001;
psychophysical improvement during the motor task
As a marker for the reproducibility of the neurophysio-
logical data we found no differences in the SOA between
Comparing the baseline performance during the first
placebo and fluoxetine prior to the motor task (placebo:
10 min we found no significant differences between placebo
(31.53 ^ 4 ms, mean ^ standard error) and fluoxetine
P ¼ 0:31). Using the ANOVA for repeated measurements
condition (29.45 ^ 2.4 ms; paired t-test: P ¼ 0:59).
with the within-subject factor ‘time £ condition’ we found
During the course of the task we found a significant
significantly increased SOA ðFð1;16Þ ¼ 13:33; P ¼ 0:002;
shortening of the latency differences between the onset of
post-hoc t-test: pre- vs. post-fluoxetine: P ¼ 0:001; post:
the APB and deltoid muscle contraction (ANOVA with
1446 ^ 168 mV, ). Under placebo we found no
within-subject factor ‘time’: placebo: Fð1;8Þ ¼ 23:93;
changes of the SOA after the completion of the task
P ¼ 0:001; fluoxetine: Fð1;8Þ ¼ 49:96; P ¼ 0:0001) indicat-
(post: 1379 ^ 127 mV, t-test: P ¼ 0:92).
ing a significant motor performance improvement underboth conditions. In the placebo session, the mean latency
during the first 10 min was 31.53 ^ 4 ms, and during the last
We found no differences in the COG between placebo
10 min 12.38 ^ 1.1 ms (t-test: P ¼ 0:001). The Bonferroni-
and fluoxetine prior to the motor task using paired t-test
(x-coordinate: P ¼ 0:25; y-coordinate: P ¼ 0:51;
differences between the first and the successive 10 min
In the placebo session, we found a significant medial shift of
(1 – 10 vs. 11 – 20: P ¼ 0:2). Afterwards subjects showed
the y-coordinate of the COG after the completion of the
increasingly reduced time differences between the onsets of
motor task (pre: 2 4.66 ^ 0.26 cm, post: 2 4.54 ^ 0.26 cm,
both muscles in the following periods that all reached
t-test: P , 0:001). We also found a significant medial shift
significant level (1 – 10 vs. 21 – 30: P ¼ 0:03; 1 – 10
vs.31 – 40: P ¼ 0:04; 1–10 vs. 41–50: P ¼ 0:01; 1–10 vs.
2 4.59 ^ 0.23 cm, t-test: P , 0:001; ANOVA, within-sub-
51 – 60: P ¼ 0:009). In the fluoxetine session we also found
ject factor ‘time £ condition’: Fð1;16Þ ¼ 24:95; P ¼ 0:0001).
significantly reduced latencies (from 29.45 ^ 2.4 ms to
Contrarily, we found no significant differences for
Fig. 1. A: Shown is the motor improvement during each session of the co-contraction task. Shown are the results obtained under placebo and fluoxetinecondition. The whiskers represent the standard error.
Table 1Sown are the individual SOA (mV), the y- (refers to the lateral-to-medial axis) and x-coordinate (refers to the posterior-to-anterior axis) of the COG (cm) and the size of the area (cm2) elicited by TMS underplacebo and fluoxetine condition
‘time £ condition’Within-subject factor
Below we show group data (mean, standard error) as well as the results of statistical analysis (ANOVA for repeated measurements, post-hoc t-test).
B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
the x-coordinate of the COG (placebo: pre: 2 0.17 ^
post-session, we found equal motor thresholds (placebo:
0.19 cm, post: 2 0.09 ^ 0.21 cm; fluoxetine: pre: 2 0.34 ^
41.6 ^ 1.9%, fluoxetine: 43.3 ^ 1.1%).
0.25 cm, post: 2 0.31 ^ 0.24 cm; ANOVA, within-subjectfactor ‘time £ condition’: Fð1;16Þ ¼ 0:52; P ¼ 0:47Þ:
In the present study we investigated the influence of the
Before the motor task we found non-significant
selective 5-HT reuptake inhibitor fluoxetine on use-depen-
differences in the size of the area between placebo and
dent plastic changes in human primary motor cortex.
fluoxetine that was also indicative for the reproducibility of
Healthy right-handed subjects performed a motor task
the motor maps (placebo: 10.7 ^ 0.78 cm2, fluoxetine:
consisting of a co-contraction of two limb muscles.
10.5 ^ 0.57 cm2, t-test: P ¼ 0:84; Afterwards
This paradigm has been proved to be an appropriate
motor maps significantly enlarged (ANOVA, within-subject
model to study the pharmacological modulation of train-
Under placebo we found a tendentious enlargement of the
motor map after the motor task (post: 11.1 ^ 0.74 cm2,
repetitive co-contraction resulted in progressively shortened
t-test pre vs. post: P ¼ 0:08). Under fluoxetine the
time intervals between the onsets of both muscles (
enlargement of the motor map, however, reached significant
level (post: 12.1 ^ 0.81 cm2, t-test pre vs. post: P ¼ 0:008).
) without significant differences between both
The shows the results of the MEP mapping performed
conditions. This lack of pharmacological effects on
immediately before and after the completion of the motor
psychophysical level was surprising as Pariente et al.
task under both conditions in one single-subject and the
previously reported a significantly improved finger
changes in motor performance during the course of the task.
tapping rate and grip force of the hemiparetic side inpost-stroke patients under a single dose of fluoxetine
We found no differences in the MT between placebo and
However, our findings are in line with previous studies
fluoxetine prior to the motor task ðP ¼ 0:31Þ: In the pre and
that also failed to show pharmacological influences on
Fig. 2. Single-subject co-contraction effect: shown are the results of the TMS mapping performed before and after the completion of the motor task under bothconditions (placebo: above; fluoxetine: below). The graphs between both motor maps show the changes in motor performance during the course of the task(placebo: from 24.9 ^ 2.3 (mean values ^ standard error) to 13.8 ^ 1.2 ms, paired t-test 0 – 10 vs. 51 – 60 min P , 0:001; fluoxetine: from 27.5 ^ 2 to15.8 ^ 1 ms; P , 0:001). Note the differences of changes in motor maps between placebo (above) and fluoxetine condition (below), especially theenlargement of the area and the gain in SOAs.
B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
fluoxetine seem therefore to have complex influences on
One hour of co-contraction resulted in significantly
different parameters of cortical excitability.
shortened time intervals between the onsets of the APB
In summary, our findings provide evidence for a
and the deltoid muscle, which was not affected by the
use-dependent facilitating effect of fluoxetine on human
administration of amphetamine. Motor improvement,
motor cortex excitability. However, we found no gain in
however, resulted in a significant medial shift of the APB
motor performance after a single dose of fluoxetine.
output map. Consequently, the co-contraction task may be
Long-term treatment may additionally improve motor
considered as an appropriate task to investigate facilitating
influences on re-organizational changes of human primary
motor cortex. But it seems not appropriate to study parallel
effects on behavioral level. In line with previous studies,
gating the influence of long-term treatment with selective
we also found a medial shift of the APB motor output map
5-HT reuptake inhibitors as an adjunct to physical therapy
under both conditions. This shift was orientated towards the
may therefore provide deeper insight into their possible
representation of the deltoid muscle. Therefore it might
therapeutical efficiency in post-stroke patients.
be indicative of a change in the underlying corticalrepresentation ().
Synchronised neuronal activity that parallels cortical
reorganization can be referred to a principle presented byHebb (He suggested
This study was supported by a grant from the
that individual neurons could participate in different cell
Ruhr-University Bochum (FORUM F 291/01) and by a
assemblies and be involved in multiple functions and
grant from the Scientific Research Council of BG-Kliniken
representations (Long-term potentiation (LTP)
Bergmannsheil, Bochum (to P.S.). We thank Mr Steve
of motor synapses can be induced in Hebbian fashion
Langan for skilful editing of the manuscript.
by pairing weak presynaptic stimulation with strongpost-synaptic depolarization (for a review see This mechanism can also function only at proximal
synapses, suggesting that dendritic spikes participategenerally in a form of synaptic potentiation that does not
Altamura AC, Moro AR, Percudani M. Clinical pharmacokinetics of
require post-synaptic action potential firing in the axon
fluoxetine. Clin Pharmacokinet 1994;26:201 – 14.
Barbeau H, Rossignol S. Initiation and modulation of the locomotor pattern
representation, which are suggested by our results might
in the adult chronic spinal cat by noradrenergic, serotonergic and
therefore depend on the synchronicity of the co-contraction
dopaminergic drugs. Brain Res 1991;546:250 – 60.
Bieger D. Role of bulbar serotonergic neurotransmission in the
movement. This in turn might induce LTP-like processes
initiation of swallowing in the rat. Neuropharmacology 1981;20:
that promote re-organizational changes of the participating
Cazalets JR, Sqalli-Houssaini Y, Clarac F. Activation of the central pattern
Several animal experiments showed purposeful motor
generators for locomotion by serotonin and excitatory amino acids in
responses due to the application of 5-HT (
neonatal rat. J Physiol 1992;455:187– 204.
Chen Y, Peng L, Zhang X, Stolzenburg JU, Hertz L. Further evidence that
fluoxetine interacts with a 5-HT2C receptor in glial cells. Brain Res
neurons were activated in association with increased muscle
motor activity, especially if the motor activity is in the
Cohen LG, Gerloff C, Ikoma K, Hallett M. Plasticity of motor cortex
repetitive or central pattern generator mode (
elicited by training of synchronous movements of hand and shoulder.
Thus, serotonergic neurons seem to promote
motor output if it is generated in Hebbian fashion.
Dam M, Tonin P, De Boni A, Pizzolato G, Casson S, Ermani M, Freo U,
Piron L, Battistin L. Effects of fluoxetine and maprotiline on functional
The present findings corroborate the hypothesis of a
recovery in post-stroke hemiplegic patients undergoing rehabilitation
reorganization processes of the APB representation. Using
Dinse HR, Ragert P, Pleger B, Schwenkreis P, Tegenthoff M. Pharmaco-
TMS, Ilic et al. previously investigated the effects of the
logical modulation of perceptual learning and associated cortical
selective 5-HT reuptake inhibitor sertaline on human motor
reorganization. Science 2003;301:91– 4.
Glanzman DL. Post-synaptic regulation of the development and long-term
cortex excitability in healthy subjects (
plasticity of Aplysia sensorimotor synapses in cell culture. J Neurobiol
Under the influence of sertaline, they found a steeper
intensity curve suggesting an increased excitability of the
Golding NL, Staff NP, Spruston N. Dendritic spikes as a mechanism for
cortico-spinal neurone. In the present study, under
cooperative long-term potentiation. Nature 2002;418:326 – 31.
the influence of fluoxetine the repetitive co-contraction of
Gram L, Fluoxetine . Engl J Med 1994;331:1354 – 61. Hebb DO. The organization of behaviour. New York: Wiley; 1949.
the APB and the deltoid muscle resulted in an increase of
Hrdina PD, Vu TB. Chronic fluoxetine treatment upregulates 5-HT uptake
the SOA and an enlargement of the APB representation.
sites and 5-HT2 receptors in rat brain: an autoradiographic study.
Selective 5-HT reuptake inhibitors like sertaline and
B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
Ilic TV, Korchounov A, Ziemann U. Complex modulation of human motor
Pleger B, Schwenkreis P, Dinse HR, Ragert P, Hoffken O, Malin J-P,
cortex excitability by the specific serotonin re-uptake inhibitor sertra-
Tegenthoff M. Pharmacological suppression of plastic changes in
line. Neurosci Lett 2002;319:116 – 20.
human primary somatosensory cortex after motor learning. Exp Brain
Jacobs BL, Fornal CA. Activity of serotonergic neurons in behaving
animals. Neuropsychopharmacology 1999;21:9S – 15S.
Pleger B, Fo¨rster A-F, Ragert P, Dinse HR, Schwenkreis P, Malin J-P,
Karson CN, Newton JE, Livingston R, Jolly JB, Cooper TB, Sprigg J,
Nicolas V, Tegenthoff M. Functional imaging of perceptual learning in
Komoroski RA. Human brain fluoxetine concentrations. J Neuropsych
human primary and secondary somatosensory cortex. Neuron 2003b;
Liepert J, Terborg C, Weiller C. Motor plasticity induced by
Schwenkreis P, Pleger B, Ho¨ffken O, Malin J, Tegenthoff M. Repetitive
synchronized thumb and foot movements. Exp Brain Res 1999;125:
training of a synchronised movement induces short-term plastic
changes in the human primary somatosensory cortex. Neurosci Lett2001;312:99– 102.
Mangan PS, Cometa AK, Friesen WO. Modulation of swimming behavior
Sommi RW, Crismon ML, Bowden CL. Fluoxetine: a serotonin-specific,
in the medicinal leech. IV. Serotonin-induced alteration of synaptic
second-generation antidepressant. Pharmacotherapy 1987;7:1 – 15.
interactions between neurons of the swim circuit. J Comp Physiol [A]
Tegenthoff M, Witscher K, Schwenkreis P, Liepert J. Pharmacological
modulation of training-induced plastic changes in human motor
Palvimaki EP, Laakso A, Kuoppamaki M, Syvalahti E, Hietala J. Up-
cortex. Electroencephalogr Clin Neurophysiol Suppl 1999;51:
regulation of beta 1-adrenergic receptors in rat brain after chronic
citalopram and fluoxetine treatments. Psychopharmacology (Berl)
Tegenthoff M, Cornelius B, Pleger B, Malin J-P, Schwenkreis P.
Amphetamine enhances training-induced motor cortex plasticity. Acta
Pariente J, Loubinoux I, Carel C, Albucher JF, Leger A, Manelfe C, Rascol
O, Chollet F. Fluoxetine modulates motor performance and cerebral
Yamazaki J, Fukuda H, Nagao T, Ono H. 5-HT2/5-HT1C receptor-
activation of patients recovering from stroke. Ann Neurol 2001;50:
mediated facilitatory action on unit activity of ventral horn cells in rat
spinal cord slices. Eur J Pharmacol 1992;220:237 – 42.
People have believed since antiquity that tiny doses of toxicants can be healthful. Now hormesis, a concept oncediscredited in scientific circles, is making a surprising comeback Sipping From a Poisoned Chalice Dioxin and its chemical cousins are amongradiation punish the body at even the small-cept of hormesis “has been taken over bythe most deadly compounds on Earth. Spikeest of doses
Allergies: Allergy Triggers There are a number of different allergy-causing substances. The most common include pollen, dust mites, mold, animal dander, insect stings, latex and certain food and medication. If you have an allergy your symptoms can range from mild eye irritation and congestion to a more severe reaction causing generalized swelling and difficulty breathing. And, if you have ast

 Clinical Neurophysiology 115 (2004) 2157–2163
Fluoxetine facilitates use-dependent excitability
Burkhard Pleger*, Peter Schwenkreis, Christian Gru¨nberg, Jean-Pierre Malin, Martin Tegenthoff
Department of Neurology, Ruhr-University Bochum, BG-Kliniken Bergmannsheil, Buerkle-de-la-Camp-Platz 1, D-44789 Bochum, Germany
Objectives: In poststroke patients, fluoxetine, a selective serotonin-reuptake inhibitor, as an adjunct to physical therapy provided a better
functional recovery from motor deficits. The aim of this study was to investigate the effect of a single dose of 20 mg fluoxetine on motorlearning and associated cortical changes in healthy right-handed subjects in order to get deeper insight into its facilitating influence on humanmotor cortex.
Clinical Neurophysiology 115 (2004) 2157–2163
Fluoxetine facilitates use-dependent excitability
Burkhard Pleger*, Peter Schwenkreis, Christian Gru¨nberg, Jean-Pierre Malin, Martin Tegenthoff
Department of Neurology, Ruhr-University Bochum, BG-Kliniken Bergmannsheil, Buerkle-de-la-Camp-Platz 1, D-44789 Bochum, Germany
Objectives: In poststroke patients, fluoxetine, a selective serotonin-reuptake inhibitor, as an adjunct to physical therapy provided a better
functional recovery from motor deficits. The aim of this study was to investigate the effect of a single dose of 20 mg fluoxetine on motorlearning and associated cortical changes in healthy right-handed subjects in order to get deeper insight into its facilitating influence on humanmotor cortex.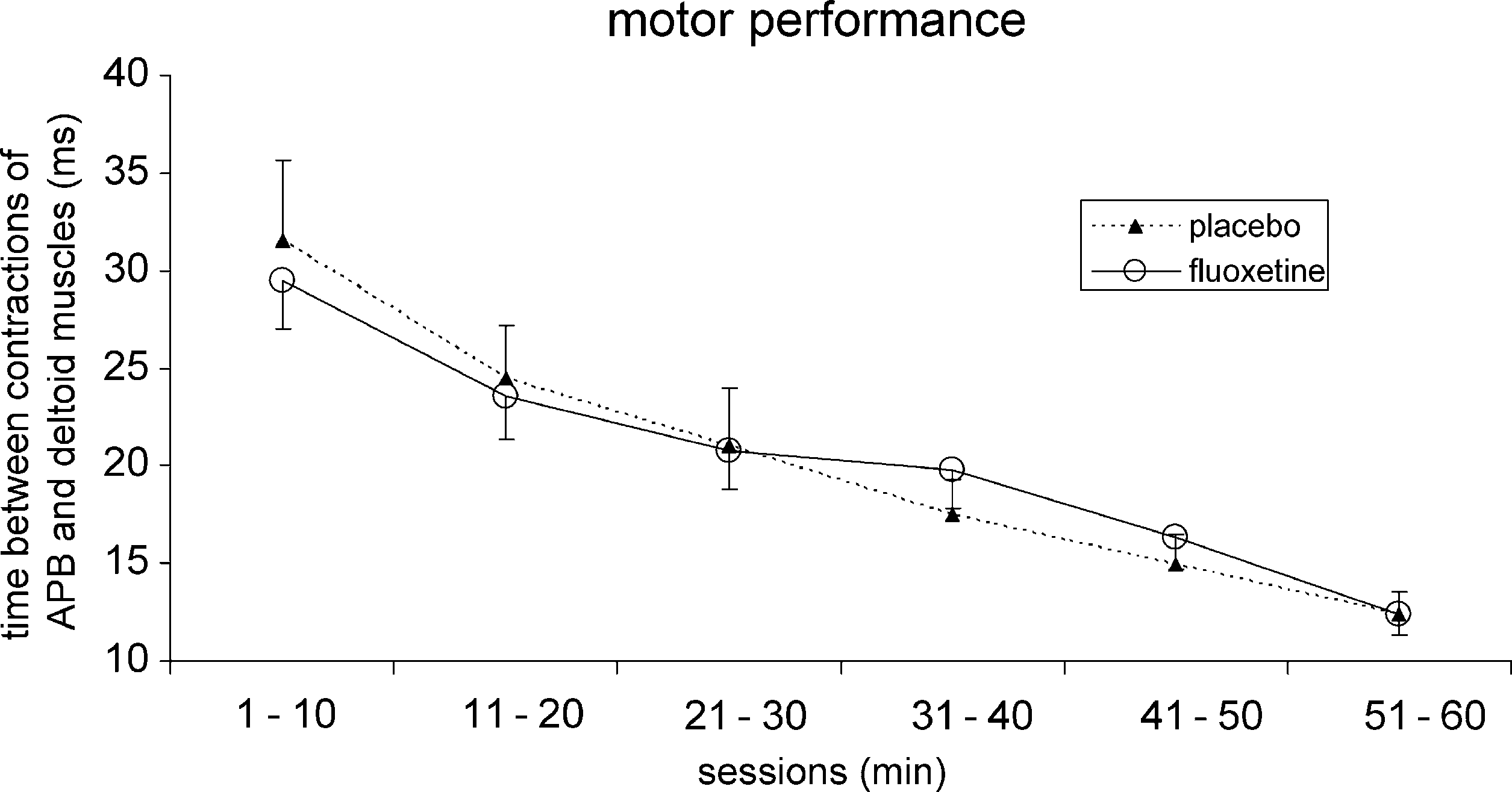 B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
by comparing the maps obtained prior to motor learning of
12.41 ^ 1 ms; P , 0:001). Using post-hoc t-test, we found
significantly reduced time differences between the first
To evaluate the effect of repetitive co-contraction on
and the successive sessions (1 – 10 vs. 11 – 20: P ¼ 0:003;
motor performance, the mean latency differences between
1 – 10 vs. 21 – 30: P ¼ 0:0003; 1–10 vs.31 –40: P ¼ 0:001;
the onsets of both muscles for the intervals 0 – 10, 10 – 20,
20 – 30, 30 – 40, 40 – 50 and 50 – 60 min were calculated in
P ¼ 0:0004). Using the ANOVA for repeated measure-
each subject and for each session. We used an ANOVA
ments and the within-subject factor ‘time £ condition’,
for repeated measurements and post-hoc t-test analysis
we found no differences between placebo and fluoxetine
(Bonferroni-corrected for multiple comparisons) to express
ðFð5;80Þ ¼ 0:59; P ¼ 0:7; ‘time’: Fð5;80Þ ¼ 39:59; P , 0:001;
psychophysical improvement during the motor task
As a marker for the reproducibility of the neurophysio-
logical data we found no differences in the SOA between
Comparing the baseline performance during the first
placebo and fluoxetine prior to the motor task (placebo:
10 min we found no significant differences between placebo
(31.53 ^ 4 ms, mean ^ standard error) and fluoxetine
P ¼ 0:31). Using the ANOVA for repeated measurements
condition (29.45 ^ 2.4 ms; paired t-test: P ¼ 0:59).
B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
by comparing the maps obtained prior to motor learning of
12.41 ^ 1 ms; P , 0:001). Using post-hoc t-test, we found
significantly reduced time differences between the first
To evaluate the effect of repetitive co-contraction on
and the successive sessions (1 – 10 vs. 11 – 20: P ¼ 0:003;
motor performance, the mean latency differences between
1 – 10 vs. 21 – 30: P ¼ 0:0003; 1–10 vs.31 –40: P ¼ 0:001;
the onsets of both muscles for the intervals 0 – 10, 10 – 20,
20 – 30, 30 – 40, 40 – 50 and 50 – 60 min were calculated in
P ¼ 0:0004). Using the ANOVA for repeated measure-
each subject and for each session. We used an ANOVA
ments and the within-subject factor ‘time £ condition’,
for repeated measurements and post-hoc t-test analysis
we found no differences between placebo and fluoxetine
(Bonferroni-corrected for multiple comparisons) to express
ðFð5;80Þ ¼ 0:59; P ¼ 0:7; ‘time’: Fð5;80Þ ¼ 39:59; P , 0:001;
psychophysical improvement during the motor task
As a marker for the reproducibility of the neurophysio-
logical data we found no differences in the SOA between
Comparing the baseline performance during the first
placebo and fluoxetine prior to the motor task (placebo:
10 min we found no significant differences between placebo
(31.53 ^ 4 ms, mean ^ standard error) and fluoxetine
P ¼ 0:31). Using the ANOVA for repeated measurements
condition (29.45 ^ 2.4 ms; paired t-test: P ¼ 0:59).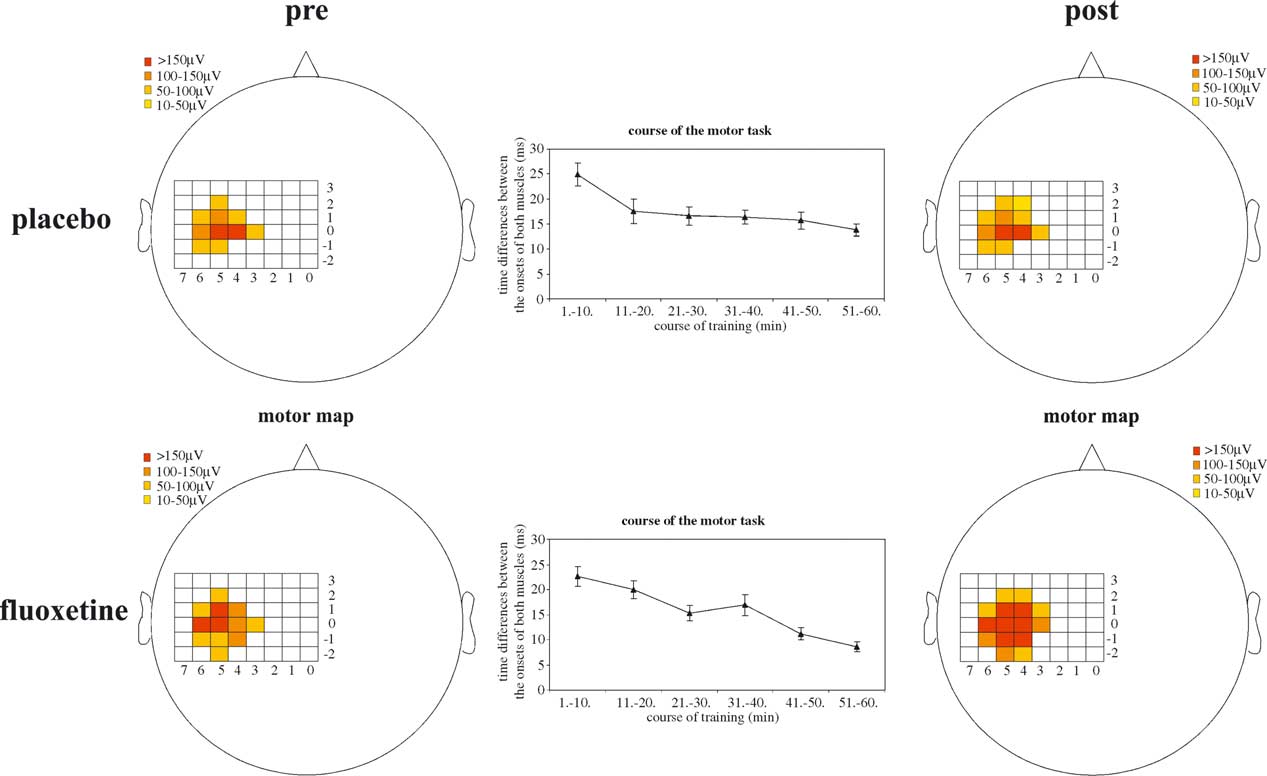 B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
the x-coordinate of the COG (placebo: pre: 2 0.17 ^
post-session, we found equal motor thresholds (placebo:
0.19 cm, post: 2 0.09 ^ 0.21 cm; fluoxetine: pre: 2 0.34 ^
41.6 ^ 1.9%, fluoxetine: 43.3 ^ 1.1%).
B. Pleger et al. / Clinical Neurophysiology 115 (2004) 2157–2163
the x-coordinate of the COG (placebo: pre: 2 0.17 ^
post-session, we found equal motor thresholds (placebo:
0.19 cm, post: 2 0.09 ^ 0.21 cm; fluoxetine: pre: 2 0.34 ^
41.6 ^ 1.9%, fluoxetine: 43.3 ^ 1.1%).