Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Jiggins.gen.cam.ac.uk
Perspectives Male-Killing Bacteria in Insects: Mechanisms, Incidence, and Implications Gregory D.D. Hurst and Francis M. Jiggins
University College London, United Kingdom
Bacteria that are vertically transmitted through female hosts and kill male hosts thatinherit them were first recorded in insects during the 1950s. Recent studies have shownthese “male-killers” to be diverse and have led to a reappraisal of the biology of manygroups of bacteria. Rickettsia, for instance, have been regarded as human pathogenstransmitted by arthropods. The finding of a male-killing Rickettsia obligately associatedwith an insect suggests that the genus’ members may be primarily associated witharthropods and are only sometimes pathogens of vertebrates. We examined both howkilling of male hosts affects the dynamics of inherited bacteria and how male-killingbacteria affect their host populations. Finally, we assessed the potential use of thesemicroorganisms in the control of insect populations.
investment in daughters at the expense of sons.
bacteria they pass on to their progeny. These
In these cases, particular host lines produce
inherited bacteria are often beneficial symbionts
female-biased sex ratios, a trait that is inherited
that play a key role in host metabolism. In many
but curable with antibiotics. We considered one
cases (e.g., the aphid symbiont Buchnera), the
class of these, the male-killing bacteria, in which
bacteria are maintained in a special host organ,
infection of a female results in the production of
the bacteriome, with the host controlling
female-biased broods because male progeny die
transmission to progeny, and show evidence of
cospeciation (1,2). In these cases, destroying thebacteria (e.g., through antibiotic treatment)
Systematics of Male-Killing Bacteria
causes a profound loss of host performance. In
Molecular systematic approaches have shown
other cases, inherited bacteria are not integrated
that male-killing bacteria derive from many
into host physiology and anatomy and do not
different clades. In most cases, the data come
show long-lived relationships with their host, as
from DNA sequencing of bacteria associated with
indicated by a lack of cospeciation (3). These
the trait and confirmation of the trait association
bacteria may be broadly separated into two
by polymerase chain reaction across infected and
classes. First, bacteria maintained through a
uninfected lines. Because inherited microorgan-
phase of horizontal transmission (e.g., Rickettsia
isms are difficult to culture, Koch’s postulates
prowazekii), with transmission to other arthro-
have been fulfilled formally in only two cases
pod hosts often occurring through a vertebrate or
(4,5). Given this caveat, male-killing bacteria
plant intermediate host (infection of the
have been found within the genus Spiroplasma
(Mollicutes) (4,6), the Flavobacteria-Bacteroides
infection follow from host feeding); second,
group (7), and the gamma and alpha subdivisions
bacteria that rarely show horizontal transmis-
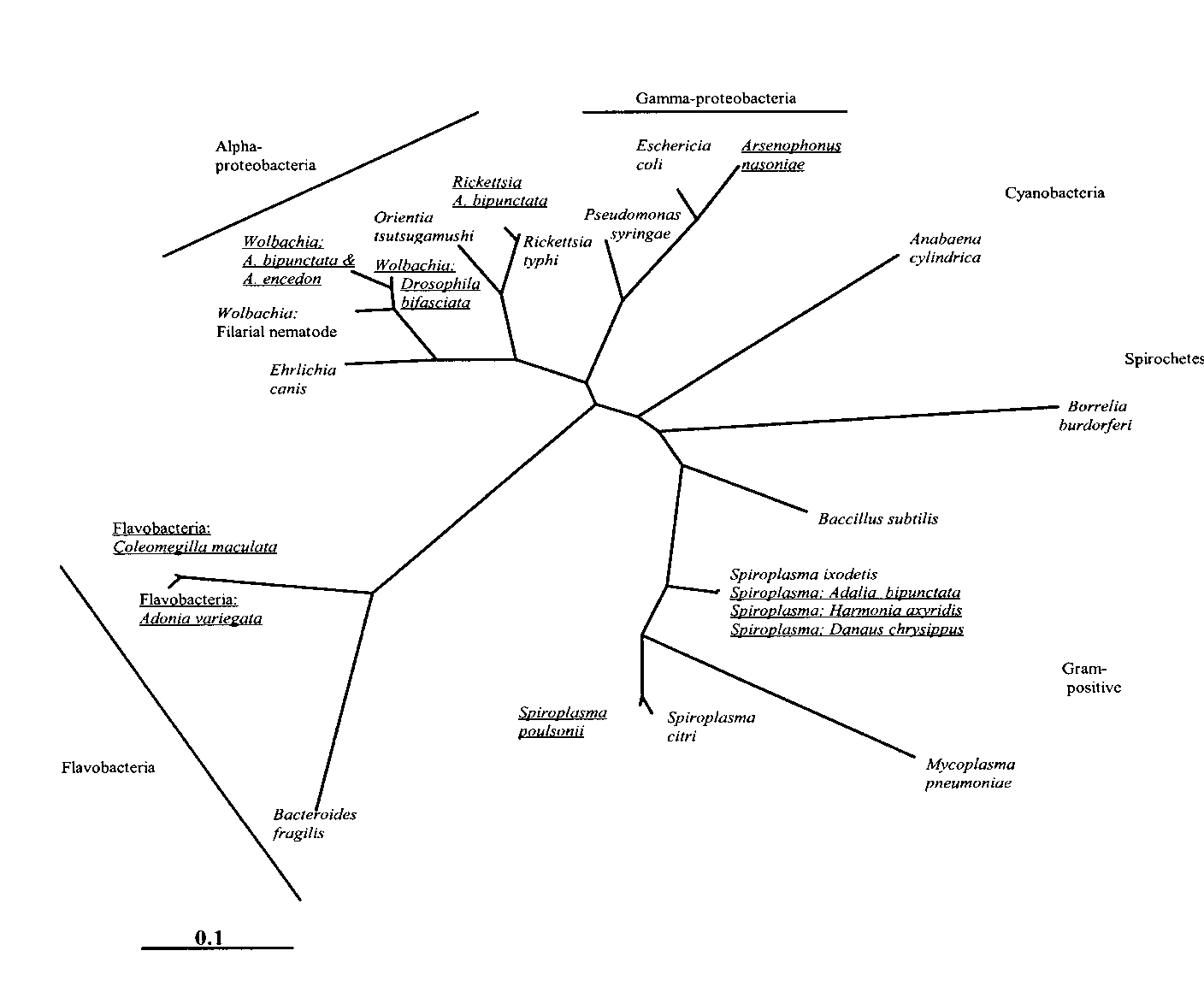
of the proteobacteria (5,8,9) (Figure).
sion, but are maintained because they manipu-
Male-killing bacteria derive from arthropod-
late host reproduction. One set of manipulations
associated bacterial clades that are not them-
manifested by these bacteria is increasing
selves male-killers. The clades can be separatedinto two types according to the transmissionmechanisms of bacteria within them: first,
Address for correspondence: Department of Biology, UniversityCollege London, 4 Stephenson Way, London NW1 2HE, U.K.;
entirely horizontal transmission or a mix of
Fax: +44 20 73832048; e-mail: g.hurst@galton.ucl.ac.uk.
horizontal and vertical transmission; and second,
Perspectives
Figure. Phylogenetic re-lationship of male-kill-ers and a selection ofother eubacteria in-ferred from 16S rDNAsequences, using maxi-mum likelihood imple-mented on PAUP*. Themale-killing bacteria(underlined) have beenlabeled with the nameof their insect host if aspecies name is notavailable. The relation-ships of the major bac-terial groups are uncer-tain.
horizontal transmission that is not epidemiologi-
established. The fact that male-killers derive
cally important. In the first type of clade are the
from such groups suggests this possibility.
genera Spiroplasma and Rickettsia. In
Spiroplasma, most members have either hori-
transmission rates far exceed those of horizontal
zontal transmission only (after feeding on a plant
transmission. Wolbachia and the flavobacterial
host) or a mix of horizontal and vertical
lineage associated with arthropods are in this
transmission between arthropod hosts (10).
group. Wolbachia are usually maintained
Rickettsia most commonly have a mix of
through manipulation of their host’s reproduc-
horizontal and vertical transmission, with
tion (12). The closest relative of the flavobacterial
horizontal transmission occurring after feeding
male-killer is Blattabacterium, the beneficial
on a vertebrate host. As recently as 10 years ago,
inherited bacterium of cockroaches and ter-
Rickettsia was regarded as one of the vertebrate
Male-killing, a trait that evolves in bacteria
Rickettsia that show transmission after feeding
already maternally inherited in arthropods, can
on plant hosts are increasingly being recognized
occur if the ancestral agent is obligately vertically
(11), and the finding of a male-killing Rickettsia
transmitted or a mix of vertical and horizontal
in ladybird beetles (8) suggests that the group is
transmission is present. Therefore, male-killing
associated with arthropods, some members of
strains are likely to be common in the genus
which cause disease in vertebrates. Other male-
Spiroplasma and the alpha group of
killing strains of Rickettsia will most likely be
proteobacteria. Furthermore, the diversity of
found. However, whether a bacterium from these
agents suggests that there is no taxonomic bar to
groups could evolve male-killing yet retain
where the transition to male-killing can take
horizontal transmission between females via
place. Thus male-killing strains are also likely to
feeding on a plant or vertebrate host has not been
be found in the spirochetes and perhaps the beta
Perspectives
and delta divisions of the proteobacteria, as these
groups are known to be vertically transmitted
coccinellid (ladybird) beetles. Approximately half
of aphidophagous species bear male-killers, and
Although vertical transmission of male-killing
one species (Adalia bipunctata) is host to at least
bacteria is the rule, transmission between host
three male-killing bacteria (6,8,9).
species has occurred. In Spiroplasma, the relatives
of S. ixodetis cause male-killing in distantly
only in insects. However, the range of insect hosts
related hosts (a butterfly and a ladybird beetle)
(6,14). The evolutionary distance between beetles
determination systems. Given that close rela-
and butterflies indicates that the bacteria do
tives of male-killing bacteria are found in
cross between host species over evolutionary time.
noninsect arthropods (e.g., Spiroplasma andRickettsia in ticks) and the conditions for the
Host Species Affected
spread of male-killing strains are met outside
The incidence of male-killing bacteria varies
insect hosts, cases of male-killing are likely to
with host ecology and biology. The trait of male-
occur in species other than insects. Two examples
killing is adaptive when the death of males
merit particular examination. First, infection
promotes the survival of female siblings. If the
with Orientia tsutsugamushi is associated with
bacteria can be transmitted only vertically, the
production of all-female broods in the trombiculid
death of male hosts can at worst be neutral (i.e.,
mite, Leptotrombidium fletcheri (18,19); in this
they cannot transmit the bacterium). Death of
example, the nature of the resultant sex-ratio
males is adaptive if it increases the survival of
distortion (primary vs. secondary bias) needs to
sibling females, who bear the same bacterium by
be assessed. Second, in the case of Spiroplasmaixodetis and its tick host Ixodes pacificus, the
The features of host biology and ecology that
association of closely related bacteria with male-
increase the benefit to the bacterium of killing
killing in insects needs to be assessed.
male embryos are sibling egg consumption(females eat their dead brothers), antagonistic
Prevalence of Male-Killers
interactions between siblings (male-killing may
in Natural Populations
reduce both cannibalism of females and the
The prevalence of male-killers in natural
intensity of competition between siblings), and
populations varies with host species (Table 1). A
deleterious inbreeding (15-17). These observa-
prevalence value of 5%-50% might be “normal”
tions explain why male-killer hosts commonly lay
among female hosts; however, in some cases
eggs in clutches. Incidence is highest where there
prevalence is very low (e.g., 1% in Drosophila
Table 1. Prevalence of male-killers in natural populations of their insect hosts (proportion of females infected)
Perspectives willistoni [20]), and in some exceptional species >
causes death in males only). What we know
90% of females are infected (e.g., the butterfly
derives almost exclusively from study of the
Acraea encedana [21]). However, there is likely to
interaction between Spiroplasma poulsonii with
be study bias towards high-prevalence infections,
and all very low-prevalence infections occur in
Studies of embryos from D. willistoni lines
drosophilids, where large samples can easily be
infected with S. poulsonii show that death occurs
bred. Infection prevalence also commonly varies
at two stages (23): 1) before gastrulation,
between populations within a host, and preva-
associated with abnormal cleavage patterns; in
lence can vary on a remarkably small scale. In the
particular, achromatic spindles, with other
walnut leaf beetle (Gastrolina depressa) in
abnormalities of the mitotic process, which
Japan, male-killers are absent in populations at
account for most embryonic deaths in male-killed
the north and south of the islands but present in
lines. 2) After gastrulation, not associated with
50%-80% of females in the center of the islands
the normal brown coloration of necrotic embryos;
(22). Prevalence variation on a kilometer scale
rather, the embryo blackens as a result of
exists in Acraea encedon (21).
breakdown of internal structures and pycnosis of
Prevalence is determined by the physiologic
effect of infection on female host performance,
The points of interaction between host and
the transmission efficiency of the bacterium from
bacterium have been investigated in D. melanogaster
mother to progeny, and the level of advantage to
lines transfected with S. poulsonii. In Drosophila,
male-killing (determined by host factors such as
sibling egg consumption) (Table 2). Transmission
chromosomes to autosomes. In females, which
efficiency may be influenced by the environment
are 2X:2n, the peptide Sxl is produced. Sxl
(e.g., high temperatures may lower transmission
induces female development of the soma and the
efficiency), the bacterium, and the host. Selection
germ line. In males, which are X: 2n, Sxl is not
favors host genes that impede the transmission of
produced. Absence of Sxl is associated with
the bacteria from mother to progeny. The spread
upregulation of genes on the single X chromo-
of host resistance genes may prevent infections
from commonly reaching the high prevalence
development, and male germ line development.
achieved by other inherited bacteria.
In Drosophila, the male-killer does not interactwith any part of the somatic sex development
Table 2. Factors affecting the prevalence of male-killing
pathway. Mutants of the tra gene bear two X
chromosomes and produce Sxl but develop as
Increase
somatic males. They are not, however, killed by
Decreased rate of inbreeding suffered by female hosts
S. poulsonii (24). Thus, the interaction between
Increased access to early resources through
male-killer and host is not associated with
somatic sex, so the target of detection and
Increased access to resources due to reduced
virulence is either before Sxl is produced, Sxl
competition, following death of sibling male hosts
itself, or the dosage compensation or germ-line
Decrease
Although the interaction between Drosophila
Inefficiency in vertical transmissionDirect physiologic costs of infection
and S. poulsonii is the only one studied in any
detail, it appears that the mechanism of sex
determination exhibited by different male-killerhosts varies widely. Male-killing bacteria havebeen observed in male heterogametic, femaleheterogametic, and haplodiploid hosts. Fur-
Mechanism of Male-Killing
thermore, members of the same clade of male-
Little is known about how male-killing is
killers can be found in hosts of different sex
achieved. Neither the cue used to detect sex nor
determination systems. The same Spiroplasma
the mechanism by which death is brought about
kills males in ladybirds (male heterogametic) and
is known in any detail. Indeed, rather than two
butterflies (male homogametic). Similarly,
steps (detection then virulence) there may be
male-killing Wolbachia have been observed in
only one (constitutive production of a factor that
both male and female heterogametic species
Perspectives
(9). Given that male and female heterogametic
Empiric studies have suggested that infec-
tion usually decreases the performance of female
directions and show different patterns of
hosts (25,26). The one exception is the interaction
dosage compensation, the fact that male-killers
between Spiroplasma poulsonii and members of
operate in both these hosts suggests that the
the Drosophila willistoni group, in which larval
development is accelerated by infection (27,28).
dosage compensation pathway may not be the
However, infection is also associated with
focus of male-killing activity; rather, somatic
increased sterility and decreased longevity
sex determination or germ-line sex determina-
among adult females (28). Male-killing bacteria,
unlike beneficial symbionts, are spread through-
Experiments with S. poulsonii demonstrate
out host tissues, and the bacteria may be present
that the somatic sex determination system is not
in very high numbers. Drosophila are infected
the focus of male-killing behavior. In the case of
with extremely high titers of S. poulsonii within
the other male-killing Spiroplasma, the presence
the hemolymph (29). Adalia bipunctata hemocytes
of the bacterium in species of different sex
are regularly infected with Rickettsia (30).
determination systems suggests that the focus is
Beneficial effects of male-killing bacteria on
either the somatic sex determination or the germ-
host performance cannot yet be ruled out.
line determination system. Two conclusions are
However, positive effects may be fewer than
therefore possible: germ-line determination is
those found in the “classical” beneficial agents,
the focus of male-killing in all cases, or male-
which typically perform a vital metabolic
killing has more than one basic mechanism.
function that insects are unable to perform. Male-
Further research is clearly warranted.
killers infect a minority of females and are rarelycarried by larval or adult males. Thus, although
Direct Effects on Female Hosts
they may add to host performance, they cannot
The interaction between male-killing bacte-
substitute for any part of it. A host cannot be
ria and their female hosts is interesting. On the
dependent on a male-killer for a physiologic
one hand, there is selection for a reduction in the
function as it can on a beneficial symbiont.
number of bacteria present in the host(minimizing virulence) and for a direct
Population and Evolutionary
physiologic contribution to host metabolism. Effects on Hosts
Invasion of a host population by male-killing
associated with the fidelity of their transmis-
bacteria affects the dynamics of the host
sion to progeny. There may be a trade-off
population and alters the pattern of selection on
between minimizing virulence and maximizing
the population to ameliorate the effects of the
vertical transmission efficiency, especially if
parasite (Table 3). A high prevalence of male-
such efficiency is positively related to bacterial
killers may increase the proportion of female
number. Thus these bacteria can be either
hosts that fail to mate (31), potentially reducing
detrimental (if the density of bacteria is high to
the population size of the host. A dearth of males
ensure vertical transmission) or beneficial to the
can subtly alter the mating system of the host.
host (if the bacteria play a role in host
Choice by females of male mates and competition
among males for mating opportunities are the
Table 3. Population and evolutionary effects of invasion of a host by male-killing bacteria
Reduced population density at larval level
due to death of male embryosFailure of females to find mates where parasite
Selection for genes that prevent transmission
prevalence leads to shortage of males, with
potential effects on adult population sizeAltered epidemiology of sexually transmitted
Alteration in host pattern of sexual selection due to
pathogens due to increased reproductive success
Perspectives
rule in insects. However, the biased population
to reduce host population size. Alternatively,
sex ratios that result from the spread of male-
killing bacteria can reverse this pattern (31).
schemes based on release of sterile males, so that
Male choice of females and competition among
they may amplify the effect of sterile releases on
females for males is expected, with a relaxation of
the population size of adult males. In addition,
selection on males to ensure paternity.
the recent discovery of male-killing in the clade
Male-killers that have invaded populations
Wolbachia adds an extra dimension to the use of
may cause changes to host biology. Theory predicts
this organism in direct and transgenic control of
selection for an increase in the size of clutch
produced (32). Most importantly, genes that
The usefulness of male-killers in reducing
prevent the action or transmission of the parasite
pest damage on their own is debatable. Insect
will be favored. The presence of these genes has
population size and population persistence are
been reported (33), but their nature and mode of
largely a function of female, not male, number.
action are unknown. The means by which insects
Thus, although the presence of a male-killer may
exclude bacteria is clearly of great import in our
reduce larval density, it is unlikely to decrease
understanding of insect-borne diseases, and the
the population size of breeding females.
nature of resistance genes is expected to be an
Furthermore, the presence of density depen-
dence during the larval stages is likely to reduce
One of the issues to be determined relates to
the effect of male death on numbers of larvae.
whether male-killing bacteria can cause the
Perhaps a more realistic use of male-killing
extinction of their host. The case of the butterflies
Acraea encedon and A. encedana is suggestive.
conjunction with sterile male release systems of
The Wolbachia male-killer in these species is at
control. In sterile male release, control is
high prevalence and clearly has some impact on
achieved through release into the environment of
the host population (21,31). If a male-killing
mass-produced sterile males, which mate with
bacterium showed perfect vertical transmission,
females and lower their fertility (34). The success
host extinction would be likely. However,
of sterile male release depends on maintaining a
selection on the host acts to lower bacterial
high ratio of sterile to normal males in the
transmission efficiency, which may ultimately
population. The presence of a male-killer in the
host population lowers the number of fertilemales and thus increases the effectiveness of any
Conclusions: Implications and Uses
release. The effects of male-killing bacteria at
of Male-Killing Bacteria
different prevalences on sterile male release, in
Male-killing is an adaptive trait that aids the
conjunction with the effects on host population
spread of inherited bacteria through natural
dynamics, need to be investigated. However,
populations. The presence of male-killing strains
direct use of male-killing bacteria as an aid to
in many bacterial taxa clearly indicates that
controlling host numbers is only achievable as a
male-killing should be considered in epidemio-
long-term stratagem. Following release of
logic investigations of vertically transmitted
infected hosts into natural populations, spread
bacteria. Male-killing is perhaps most important
will occur only in hosts with suitable ecologies
in interactions between arthropods and Rickett-
and significant prevalence levels will be achieved
sia and Spiroplasma. Members of these genera
over a period of years rather than weeks.
frequently show horizontal transmission be-
Another potential application of male-killing
tween arthropod hosts (after host-feeding), as
bacteria in the sphere of pest and disease vector
well as vertical transmission in the arthropod
control may occur indirectly through study of the
host. Given that some bacteria in these groups
virulence mechanisms of male-killers.
induce male-killing, testing for the presence orabsence of this trait should be a part of future
Acknowledgments
investigations of their epidemiology.
The authors thank Andrew Pomiankowski and two
anonymous reviewers for their comments on the manuscript.
bacteria in pest control has yet to be properly
In conducting this study, Greg Hurst was supported by
assessed. Male-killers may be used on their own
a BBSRC D Phillips Fellowship and Frank Jiggins by aBBSRC studentship. Perspectives
Dr. Hurst has been a BBSRC David Phillips Fellow
16. Werren JH. The coevolution of autosomal and cytoplasmic
at University College London since 1997. His research
sex ratio factors. J Theor Biol 1987;124:317-34.
interests center on the dynamics and importance of para-
17. Hurst GDD, Majerus MEN. Why do maternally
sites that affect insect reproduction.
inherited microorganisms kill males? Heredity1993;71:81-95.
18. Roberts LW, Rapmund G, Cadigan FCJ. Sex ratio in
References Rickettsia tsutsugamushi infected and non-infected
1. Moran N, Baumann P. Phylogenetics of cytoplasmically
colonies of Leptotrombidium (Acari: Trombiculidae). J
inherited microorganisms of arthropods. Trends Ecol
19. Takahashi M, Urakami H, Yoshida Y, Furuya Y,
2. Baumann P, Lai C-Y, Baumann L, Rouhbakhsh D,
Misumi H, Hori E, et al. Occurrence of high ratio of
Moran NA, Clark MA. Mutualistic associations of
males after introduction of minocycline in a colony of
aphids and prokaryotes: biology of the genus Buchnera. Leptotrombidium fletcheri infected with Orientiatsutsugamushi. Eur J Epidemiol 1997;13:79-86.
3. O’Neill S, Giordano R, Colbert AME, Karr TL,
20. Williamson DL, Poulson DF. Sex ratio organisms
Robertson HM. 16S rRNA phylogenetic analysis of the
(Spiroplasmas) of Drosophila. In: Whitcomb RF, Tully
bacterial endosymbionts associated with cytoplasmic
JG, editors. Sex ratio organisms (Spiroplasmas) of
incompatability in insects. Proc Natl Acad Sci U S A
Drosophila; Vol. III. New York: Academic Press; 1979.
4. Hackett KJ, Lynn DE, Williamson DL, Ginsberg AS,
21. Jiggins FM, Hurst GDD, Dolman CE, Majerus MEN. High
Whitcomb RF. Cultivation of the Drosophila
prevalence of male-killing Wolbachia in the butterfly
spiroplasma. Science 1986;232:1253-5. Acraea encedana. J Evol Biol 2000;13:495-501.
5. Werren JH, Skinner SW, Huger AM. Male-killing
22. Chang KS, Shiraishi T, Nakasuji F, Morimoto N.
bacteria in a parasitic wasp. Science 1986;231:990-2.
Abnormal sex ratio condition in the walnut leaf beetle,
6. Hurst GDD, vd Schulenburg HG, Majerus TMO,
Gastrolina depressa (Coleoptera: Chrysomelidae). Appl
Bertrand D, Zakharov IA, Baungaard J, et al. Invasion
of one insect species, Adalia bipunctata, by two
23. Counce SJ, Poulson DF. Developmental effects of the
different male-killing bacteria. Insect Molec Biol
sex-ratio agent in embryos of Drosophila willistoni. J
7. Hurst GDD, Hammarton TC, Majerus TMO, Bertrand
24. Sakaguchi B, Poulson DF. Interspecific transfer of the
D, Bandi C, Majerus MEN. Close relationship of the
“sex-ratio” condition from Drosophila willistoni to D.
inherited parasite of the ladybird, Coleomegillamelanogaster. Genetics 1963;48:841-61. maculata, to Blattabacterium, the beneficial symbiont
25. Hurst GDD, Purvis EL, Sloggett JJ, Majerus MEN. The
of the cockroach. Genet Res 1997;70:1-6.
effect of infection with male-killing Rickettsia on the
8. Werren JH, Hurst GDD, Zhang W, Breeuwer JAJ,
demography of female Adalia bipunctata L. (two spot
Stouthamer R, Majerus MEN. Rickettsial relative
associated with male killing in the ladybird beetle
26. Ikeda H. The cytoplasmically inherited ‘sex-ratio’
(Adalia bipunctata). J Bacteriol 1994;176:388-94.
condition in natural and experimental populations of
9. Hurst GDD, Jiggins FM, vd Schulenburg JHG,
Drosophila bifasciata. Genetics 1970;65:311-33.
Bertrand D, West SA, Goriacheva II, et al. Male-killing
27. Malogolowkin-Cohen C, Rodriguez-Pereira MAQ. Wolbachia in two species of insect. Proc R Soc Lond B
Sexual drive of normal and SR flies of Drosophilanebulosa. Evolution 1975;29:579-80.
10. Whitcomb RF. The genus Spiroplasma. Annu Rev
28. Ebbert M. The interaction phenotype in the Drosophilawillistoni–spiroplasma symbiosis. Evolution
11. Davis MJ, Ying Z, Brunner BR, Pantoja A, Ferwerda
FH. Rickettsial relative associated with papaya bunchy
29. Sakaguchi B, Poulson DF. Distribution of “sex-ratio”
top disease. Curr Microbiol 1998;36:80-4.
agent in tissues of Drosophila willistoni. Genetics
12. Stouthamer R, Breeuwer JAJ, Hurst GDD. Wolbachiapipientis: microbial manipulator of arthropod
30. Hurst GDD, Walker LE, Majerus MEN. Bacterial
reproduction. Annu Rev Microbiol 1999;53:71-102.
infections of hemocytes associated with the maternally
13. Bandi C, Damiani G, Magrassi L, Grigolo A, Fani R,
inherited male-killing trait in British populations of the
Sacchi L. Flavobacteria as intracellular symbionts in
two spot ladybird, Adalia bipunctata. J Invertebr
cockroaches. Proc R Soc Lond B Biol Sci 1994;257:43-8.
14. Jiggins FM, Hurst GDD, Jiggins CD, vd Schulenburg
31. Jiggins FM, Hurst GDD, Majerus MEN. Sex ratio
JHG, Majerus MEN. The butterfly Danaus chrysippus
distorting Wolbachia causes sex role reversal in its
is infected by a male-killing Spiroplasma bacterium.
butterfly host. Proc R Soc Lond B Bio Sci 2000;267:69-73.
32. Hurst GDD, McVean GAT. Parasitic male-killing
15. Skinner SW. Son-killer: a third extrachromosomal
bacteria and the evolution of clutch size. Ecol Entomol
factor affecting sex ratios in the parasitoid wasp
Nasonia vitripennis. Genetics 1985;109:745-54. Perspectives
33. Cavalcanti AGL, Falcao DN, Castro LE. “Sex-ratio” in
39. Hurst GDD, Majerus MEN, Walker LE. The
Drosophila prosaltans—a character due to interaction
importance of cytoplasmic male killing elements in
between nuclear genes and cytoplasmic factors. Am
natural populations of the two spot ladybird, Adaliabipunctata (Linnaeus) (Coleoptera: Coccinellidae). Biol
34. Robinson AS. Sex ratio manipulation in relation to
insect pest control. Annu Rev Genet 1983;17:191-214.
40. Geier PW, Briese DT, Lewis T. The light brown apple
35. Majerus TMO, Majerus MEN, Knowles B, Wheeler J,
moth Epiphyas postvittana (Walker). 2. Uneven sex
Bertrand D, Kuznetsov VN, et al. Extreme variation in
ratios and a condition contributing to them in the field.
the prevalence of inherited male-killing microorganisms
between three populations of Harmonia axyridis
41. Clarke C, Sheppard PM, Scali V. All female broods in
(Coleoptera: Coccinellidae). Heredity 1998;81:683-91.
the butterfly Hypolimnas bolina (L.). Proc Roy Soc
36. Jiggins FM, Hurst GDD, Majerus MEN. Sex ratio
distortion in Acraea encedon (Lepidoptera:
42. Brimacombe LC. All-female broods in field and
Nymphalidae) is caused by a male-killing bacterium.
laboratory populations of the Egyptian cotton
leafworm, Spodoptera littoralis (Boisduval) (Lepi-
37. Hurst GDD, Hammarton TC, Obrycki JJ, Majerus TM,
doptera: Noctuidae). Bull Entomol Res 1980;70:475-81.
Walker LE, Bertrand D, et al. Male-killing bacteria in a
43. Higashiru Y, Ishihara M, Schaefer PW. Sex ratio
fifth ladybird beetle, Coleomegilla maculata (Coleoptera:
distortion and severe inbreeding depression in the
Coccinellidae). Heredity 1996;77:177-85.
gypsy moth Lymantria dispar L. in Hokkaido, Japan.
38. Hurst GDD, Bandi C, Sacchi L, Cochrane A, Bertrand
D, Karaca I, et al. Adonia variegata (Coleoptera:Coccinellidae) bears maternally inherited Flavobacteriathat kill males only. Parasitology 1999;118;125-34.
Menopause: The Journal of The North American Menopause SocietyVol. 19, No. 9, pp. 949 / 951DOI: 10.1097/gme.0b013e3182651d8d* 2012 by The North American Menopause SocietyDepression and heavy bleeding during the menopausal transition:adjusting our gazeResearchpublishedduringthepastthreedecadeshas every1to2hours.Abnormalbleedingwasdefinedasmenstrualexplored the relationship between mood and mens
I. GENERAL All Physicians and Practitioners All Specialties Section 1. Admission of Patients: Medical Staff members of the Hospital may admit patients for diagnostic, surgical and therapeutic care in accordance with their granted privileges. Exceptions are patients with serious burns and acute psychiatric diagnoses requiring locked facilities. 1.1 Admitting Physician
 Perspectives
Perspectives