Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Lgf.ib.unicamp.br
Assessing the impact of Cry1Ab-expressing corn pollen on monarch butterfly larvae in field studies Diane E. Stanley-Horn*†, Galen P. Dively‡, Richard L. Hellmich§, Heather R. Mattila*, Mark K. Sears*, Robyn Rose‡, Laura C. H. Jesse¶, John E. Loseyʈ, John J. Obrycki¶, and Les Lewis§
*Department of Environmental Biology, University of Guelph, Guelph, ON, Canada N1G 2W1; ‡Department of Entomology, University of Maryland, College
Park, MD 20742; §United States Department of Agriculture, Agricultural Research Station, Corn Insects and Plant Genetics Research Unit, ¶Department of
Entomology, Iowa State University, Ames, IA 50011-3140; and ʈDepartment of Entomology, Cornell University, Ithaca, NY 14853
Edited by M. R. Berenbaum, University of Illinois at Urbana-Champaign, Urbana, IL, and approved August 22, 2001 (received for review June 4, 2001)
Survival and growth of monarch larvae, Danaus plexippus (L.),
mine the impact of Bt pollen on the survival and growth of D.after exposure to either Cry1Ab-expressing pollen from three plexippus. Each study varied in experimental design and treat-
Bacillus thuringiensis (Bt) corn (Zea mays L.) events differing in
ments tested; however, the general approach was to expose
toxin expression or to the insecticide, -cyhalothrin, were exam-
larvae to milkweed leaves containing natural deposits of Bt and
ined in field studies. First instars exposed to low doses (Ϸ22 grains
non-Bt pollen that accounted for pollen accumulation, natural
per cm2) of event-176 pollen gained 18% less weight than those
degradation of toxin within pollen of various ages under differ-
exposed to Bt11 or Mon810 pollen after a 5-day exposure period.
ent environmental conditions, and the possible ingestion of
Larvae exposed to 67 pollen grains per cm2 on milkweed leaves Bt-expressing plant material other than pollen. In the Maryland
from within an event-176 field exhibited 60% lower survivorship
study, the responses of larvae to pollen were compared with
and 42% less weight gain compared with those exposed to leaves
responses to an insecticide used to control Ostrinia nubilalis. from outside the field. In contrast, Bt11 pollen had no effect on growth to adulthood or survival of first or third instars exposed for 5 days to Ϸ55 and 97 pollen grains per cm2, respectively. Similarly,
Iowa Study I. A split-split plot experiment was conducted on two
no differences in larval survivorship were observed after a 4-day
Iowa State University farms planted on May 24, 2000 with six Btexposure period to leaves with 504 –586 (within fields) or 18 –22
hybrids: NK3030Bt, NK7070Bt (Syngenta Seeds, Golden Valley,
(outside the field) pollen grains per cm2 collected from Bt11 and
MN; Bt11 event), 38G17, 34R07 (Pioneer Hi-Bred, Des Moines,
non-Bt sweet-corn fields. However, survivorship and weight gain
IA; Mon810 event), Maximizer 21 (Syngenta Seeds; event 176),
were drastically reduced in non-Bt fields treated with -cyhalo-
and 2249 (Mycogen Seeds, Indianapolis, IN; event 176). The
thrin. The effects of Bt11 and Mon810 pollen on the survivorship
non-Bt hybrids NK3030, NK7070 (Syngenta), 3489, and 3893
of larvae feeding 14 to 22 days on milkweeds in fields were
(Pioneer Hi-Bred) were included as controls. The transforma-
negligible. Further studies should examine the lifetime and repro-
tion events were randomly assigned as main-plot treatments to
ductive impact of Bt11 and Mon810 pollen on monarchs after
four complete blocks on each farm. Plots measured 4.6 by 4.6 m
long-term exposure to naturally deposited pollen.
and were separated by Ն18.3 m of soybeans to minimize pollen
drift among plots (9). Subplots were potted A. syriaca plants
Corn(ZeamaysL.)hasbeentransformedwithagenefromthe placed at two locations per plot Ϸ2 days before corn anthesis,
bacterium Bacillus thuringiensis (Bt) to express the insecti-
with two in the middle and two at the edge of the plot. When
cidal 1 epidopteran-active crystalline protein (Cry1Ab) endo-
50–75% of the corn plants had shed pollen, three first-instar
toxin (1). The Cry1Ab toxin is specifically active on the lepidop-
monarch larvae (Ͻ24-h old) were transferred to leaves on the
teran species so the impact on nontarget organisms has been
upper half of each milkweed plant. One plant from each subplot
considered negligible (2). However, most commercial Bt corn
was caged with fine-mesh screening to deter predation. Before
hybrids express the toxin in the pollen (3) that may be deposited
larval infestation, a leaf was selected randomly from the upper
on host plants of nontarget species. In particular, the monarch
half of each plant and brought back to the laboratory where
butterfly, Danaus plexippus (L.) may be at risk (4, 5), because
pollen grains were counted in five randomly chosen 0.25-cm2
monarch larvae feed on the common milkweed, Asclepias syriaca
sections of the leaf by using an ocular grid and a stereo
(6), in and near cornfields. About half of the overwintering
microscope. Five days after infestation, larval survival and
monarch population originates from the region known as the
weight were recorded. Each leaf then was removed from the
Corn Belt (7), so a portion of the migratory monarch population
upper half of each plant for a second determination of pollen
may be exposed to Bt pollen. However, the exposure of monarch
larvae to the Cry1Ab toxin varies for different Bt corn events.
Pollen from event-176 Bt hybrids expresses the highest level of
Ontario Study. The study was conducted in six commercial field-
corn sites [4.9–17.4 hectares (ha)] planted within Wellington
g͞g of pollen; http:͞͞www.epa.gov͞
pesticides͞biopesticides͞factsheets͞fs006458t.htm) and has
County, Ontario, Canada in May 2000. Three Bt11 fields (two
been demonstrated to have adverse effects on first-instar mon-
N2555 and one N27 M3, Syngenta Seeds) and three non-Bt fields
arch caterpillars (5, 8). The registration of hybrids derived from
(Pride 177, Pioneer Hi-Bred; Hyland 2240, Hyland Seeds, Blen-
event 176 will terminate in 2001 (http:͞͞www.epa.gov͞scipoly͞
sap͞2000͞october͞brad1 execsum overview.pdf). Bt11 and
This paper was submitted directly (Track II) to the PNAS office.
(http:͞͞www.epa.gov͞scipoly͞sap͞2000͞october͞), so the po-
Abbreviations: Bt (Bacillus thuringiensis); Cry1Ab, 1 epidopteran-active crystalline protein;ha, hectares.
tential negative impacts of these hybrids may be lower than that
†To whom reprint requests should be addressed. E-mail: destanle@uoguelph.ca.
Reported herein are five independent field studies conducted
The publication costs of this article were defrayed in part by page charge payment. Thisarticle must therefore be hereby marked “advertisement” in accordance with 18 U.S.C.
in Iowa, Maryland, Ontario (Canada), and New York to deter-
§1734 solely to indicate this fact.
www.pnas.org͞cgi͞doi͞10.1073͞pnas.211277798
PNAS ͉ October 9, 2001 ͉ vol. 98 ͉ no. 21 ͉ 11931–11936
heim, ON, Canada; and 4064NK, Syngenta Seeds) were selected
between hybrid treatments, and each block was surrounded by a
based on whether they were Ͼ150 m from other cornfields and
9-m buffer zone of bare ground to minimize drift effects. Plants
had naturally occurring milkweed in the following locations: Ϸ1
were propagated and transplanted as described above. Two
m inside the field, Ͻ1 m from the field edge, and Ϸ5 m from the
plants were transplanted at 3 m from the inside edge (between
field edge, along eight transects perpendicular to the field edge.
rows 4 and 5) and the outside edge of each hybrid plot, along
As controls, eight naturally occurring milkweed plants were
transects evenly spaced along the field edge. The air-blast
selected in each of three natural areas situated Ͼ150 m from any
sprayer ran between the edge of the plot and the outside
milkweeds, so these plants were not exposed to the directed
The eight transects were alternately assigned to one of two
spray. Separate bioassays were conducted at 3, 6, and 9 days after
bioassays that began on day 6 and day 11 of anthesis, respectively.
the onset of anthesis, as described above. Leaves were removed
For each bioassay, a cohort of five larvae, reared on A. syriaca
within 1 h after insecticide treatments that coincided with the
in the laboratory, was weighed, randomly assigned to a plant, and
placed on the upper third of the plant. The whole plant was then
enclosed in a fine mesh cage to deter predation. Larvae for the
Iowa Study II. A replicate field site subdivided into paired 0.2-ha
first bioassay were first instars (12- to 36-h old), whereas those
plots of N4640Bt corn (Syngenta Seeds; Bt11 event) and non-Bt
used in the second bioassay were third instars (Ͻ24 h post second
N4640 corn (Syngenta Seeds) was planted in late April 2000 at
molt). After a 5-day exposure period, the number and instar of
each of three Iowa State University farms. Milkweed plants were
larvae were recorded. Larval cohorts were brought back to the
transplanted to the field sites during May and June. Six plants
laboratory where they were weighed and reared to adulthood on
(5–50 cm tall) were established at each of three locations: 4.6 m
clean A. syriaca at 24°C and a 16-h light͞8-h dark cycle. Addi-
inside the plot, at the field edge, and 2 m from the plot edge. Five
tional data were recorded on days to pupation, pupal weight,
first instars were placed on each plant Ϸ7 days after the start of
percentage of pupation and emergence, and adult weight and
anthesis. The number and life stages of larvae were recorded
wing length. Leaves with feeding damage were collected from
every 24 h for the first 7 days, and then every 48 h for another
plants of the first bioassay and viewed with a video imaging
7 days. Leaves were collected and pollen densities were esti-
system (XC-75CE, Cosmicar͞Pentax 16-mm camera with
mated at 3–8 days after larval introduction by counting the
NORTHERN EXPOSURE V.2.9E software, Empix Imaging, Missis-
number of grains on one 0.5-cm2 leaf disk excised with a #6 cork
sauga, ON, Canada) to estimate the area of consumption. Pollen
borer from a middle leaf of each plant (5).
densities were determined for leaves collected from the tops,
middles, and bottoms of plants. To minimize the loss of pollen,
New York Study. One 0.25-ha field near Ithaca, NY was planted
all sampled leaves were sandwiched between strips of contact
in two 36-row sections, one with the Bt hybrid 36-G32Bt (Pioneer
paper (ConTact7 Brand; Decora Manufacturing, North Rid-
HiBred, Mon810 event) and the other with a non-Bt corn hybrid
geville, OH). Pollen counts within five 1-cm2 areas on the tops
(3752). Ten milkweed plants were transplanted at three locations
and bottoms of leaves were added to corresponding counts in the
in each section: along the edge, and at 6.6 and 32.7 m inside the
contact paper to obtain density estimates. Grains adhering to the
field. At Ϸ2 days from the onset of pollen shed, five first instars
paper were stained with acid fuchsin (Sigma) to facilitate
were placed on each plant. The number and life stages of larvae
were recorded every 24 h for 22 days. The pollen density on each
plant was measured 6 days after larval introduction according to
Maryland Study. Field-corn experiment. The study was conducted
the same methods used in the Iowa II study.
in an 8-ha cornfield at the University of Maryland Research and
Education Center at Beltsville, MD. The field was planted on
Data Analysis. In the Maryland study, variances of data not
May 3, 2000 with the hybrid TM5114 (Mycogen Seeds; event
meeting the assumptions of ANOVA were grouped before
176). Milkweed plants were propagated from rhizomes in 20-cm
analysis (10). In the Ontario and Iowa I studies, log or arcsine
pots and grown outdoors until they were 50–60 cm in height.
transformations were applied as required. In all three experi-
Two transects of milkweeds treated as blocks were established
ments, the following variables for each bioassay were analyzed by
along each side of the field before anthesis. At each transect,
ANOVA (PROC MIXED; ref. 11): pollen deposition, percentage of
plants were transplanted within and outside the field at Ϫ10, Ϫ5,
survival and weight gain, and in the Ontario study, development,
Ϫ3, Ϫ1 m, and 1, 3, 5, and 10 m from the field edge, respectively.
consumption, days to pupation, pupal and adult weights, percent
Separate bioassays with field-collected leaves from each plant
emergence, and adult wing length. In all studies, corn hybrid
position were conducted at 3, 6, 9, and 14 days after the onset of
treatment and milkweed position were treated as fixed effects,
anthesis. At each time, pollen densities were estimated by
whereas block or field was treated as a random effect. For the
counting the number of grains in a 0.34-cm2 viewing area on a
Iowa I data, bioassay type (caged or uncaged) was treated as a
single leaf that was removed from the upper one-third of each
fixed subplot factor. In addition, orthogonal contrasts were
milkweed plant. Each leaf then was infested with a cohort of 10
conducted to test for differences between each Bt11 and Mon810
first instars (Ϸ24-h old) for 4 days. Survival and weights of larval
hybrid and their isolines, and between the event-176 hybrids vs.
cohorts were assessed both initially and after 4 days.
the other two Bt hybrids combined. Means were separated with
Sweet-corn experiment. The study was conducted at the Uni-
Tukey’s studentized range test (P Ͻ 0.05). For the Iowa II and
versity of Maryland Research and Education Center at Upper
New York studies, the number of larvae alive over time was
Marlboro, MD. The plot was planted on May 12, 2000 and
analyzed by ANOVA (PROC GLM). Survival curves for larvae
consisted of three hybrid treatments: untreated Attribute
were analyzed separately for each hybrid with LIFETEST (SAS
GSS0937 (Bt), untreated Prime Plus (non-Bt isoline), and treated
Prime Plus, replicated four times in a randomized block design.
The Attribute hybrid (Syngenta Seeds) was developed by cross-
ing with the Cry1Ab (Bt11 event) field corn and then backcross-
Iowa Study I. Average pollen densities ranged from 21 to 23 grains
ing to produce a pure inbred of Bt sweet corn. The treated non-Bt
per cm2 across hybrid treatments, which were not significantly
plots received three applications of -cyhalothrin (Warrior 1E,
different. These levels were relatively low compared with pub-
Syngenta Crop Protection) delivered at the 94.6-ml formulation
lished reports of pollen deposition (12) that could be a small-plot
rate by an air-blast sprayer in 536 liters of spray volume per ha.
effect. Only the plant location effect was significant (P ϭ 0.027),
Each of the four replicate blocks were planted with 4-m buffers
showing higher pollen levels in the middle of the plot (26 grains
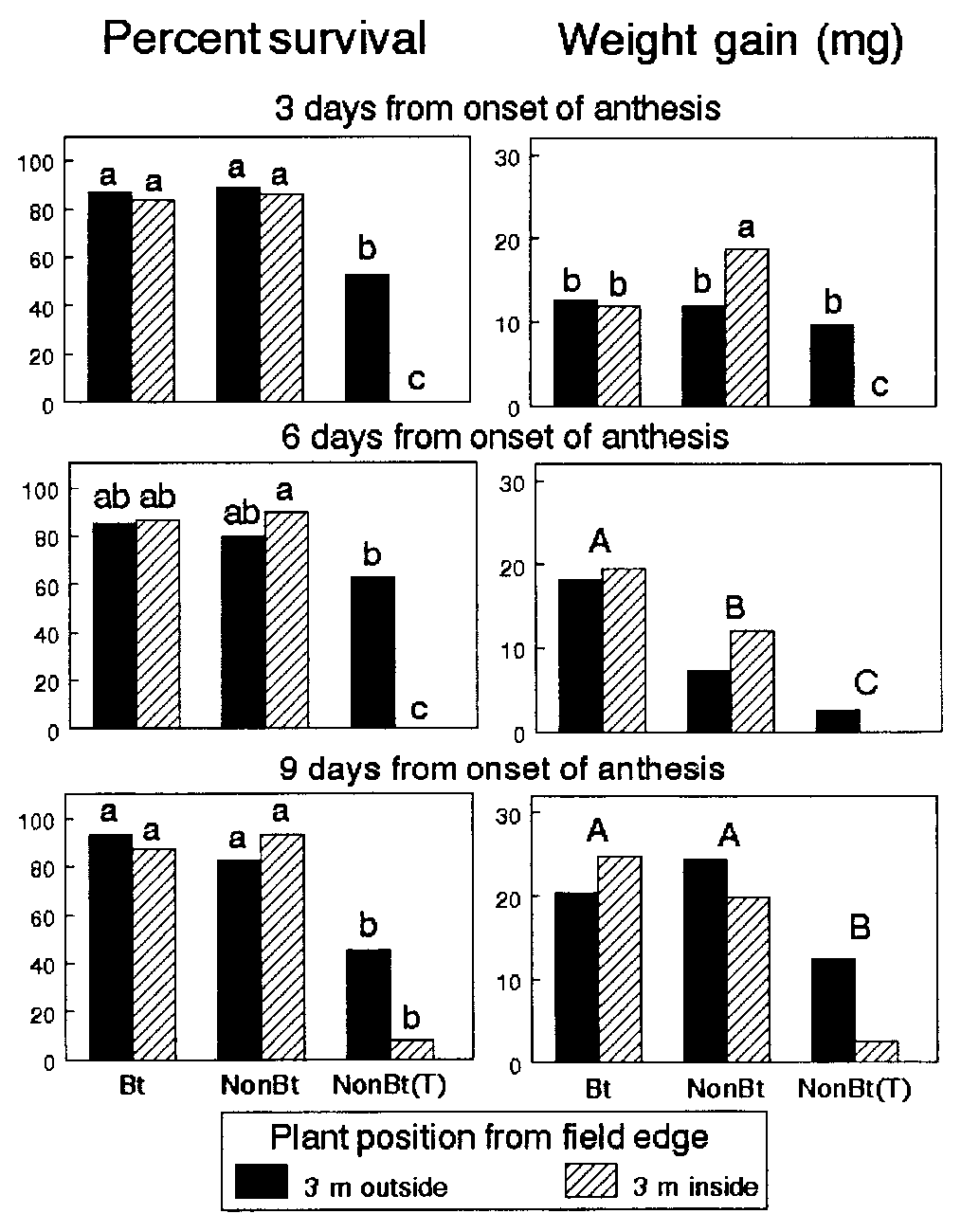
11932 ͉ www.pnas.org͞cgi͞doi͞10.1073͞pnas.211277798 Table 1. Comparison of survivorship and growth for Monarch larvae exposed as first or third instars to Bt11 and non-Bt pollen on caged milkweed (A. syriaca) plants in Ontario
*Day 5 avg instar͞day 0 avg instar. †Day 6 avg wt͞day 0 avg wt. ‡Days to pupation from the beginning of the exposure period.
per cm2) compared with the plot edge (18 grains per cm2). For
anthesis were 8.2, 63.7, 161.2, and 27.8 grains per cm2, respec-
caged tests, average weights ranged from 2.8 to 3.3 mg per larva,
tively. Average levels outside the field peaked at 18.5 grains per
and all main and interaction effects were nonsignificant. For the
cm2 on day 9 and significantly dropped off along positions
uncaged tests, no significant effects also were indicated by using
extending away from the field edge. The relative differences in
the general linear model; however, the contrast test showed that
pollen deposition across transect positions on each date were
the average weights of larvae (1.7 mg) that fed on leaves with
not the same (P ϭ 0.0015). During anthesis, seven rain events
event-176 pollen were significantly less than weights of larvae
(11–25 mm per day) occurred that may have washed pollen off
(2.0 mg) on the other Bt hybrids combined (P ϭ 0.037). For
percent survival, the hybrid treatment and all interaction effects
Survival of monarch larvae ranged from 57.5 to 86.3% on day
with hybrid were not statistically significant. However, both the
3 of anthesis (Fig. 1). Survivors, which initially weighed an
plant location (P ϭ 0.002) and bioassay-type (caged or uncaged;
P Ͻ 0.001) effects were significant. Survival was significantly
higher at the plot edge (53.6%) and in caged cohorts (61.2%),
compared with the middle of the plot (38.1%) and uncaged
cohorts (30.5%), respectively. Predatory insects may have used
the plots as a refuge to escape the prevailing hot-dry conditions
in Iowa, resulting in higher predation in the middle of the field
compared with the soy buffers surrounding the field. Also, lower
predation seemed to occur in caged cohorts.
Ontario Study. Pollen densities on leaves did not differ between
hybrid types (Bt or non-Bt) but decreased significantly with
increasing distance from the field on day 6 (P Ͻ 0.0001) and day
11 (P Ͻ 0.0001) of anthesis. On day 6, the ranges of pollen
densities at each distance were 2–309, 0–176, and 0–75 grains per
cm2 for plants found at Ϫ1 m within the field, Ͻ1 m outside the
field, and 5 m outside the field, respectively. On day 11, the range
of densities calculated on leaf samples taken from Ϫ1, Ͻ1, and
5 m were 3–429, Ͻ1–320, and Ͻ1–50, respectively. Control plants
contained Ϸ1 grain per cm2 on both days, probably resulting
from contamination during leaf sampling. Average accumulated
rainfall during the first 6 and 11 days of pollen shed (before
placing cages on plants) was Ϸ10 and 13 mm, respectively, and
may have washed pollen off of the plants (12).
The comparison of responses to Bt and non-Bt pollen is
presented in Table 1. Neither hybrid type nor plant position
affected survivorship, developmental rate, or weight gain of first-
or third-instar monarchs during the exposure period in the field
or in later developmental stages (but see % emergence for first
instars in Table 1). No significant differences in survivorship,
weight gain, or development to adulthood were observed be-
tween larvae in cages within the fields and those in control cagesϾ150 m from any cornfield in either bioassay (P Ն 0.15 for all
Survival of first-instar monarch larvae feeding on milkweeds placed
at 1, 3, 5, and 10 m inside (negative values) and outside the field edge ofevent-176 field corn in Maryland. Means Ϯ SE are based on separate bioassays
Maryland Study. Field-corn experiment. The average pollen den-
conducted on days 3, 6, 9, and 14 of anthesis. Within each graph, columns with
sities on milkweed leaves in the field on days 3, 6, 9, and 14 of
the same letters are not significantly different (P Ͻ 0.05; Tukey’s test).
PNAS ͉ October 9, 2001 ͉ vol. 98 ͉ no. 21 ͉ 11933
Survival and weight gain of first-instar monarch larvae feeding on
Weight gain of first-instar monarch larvae feeding on milkweeds
milkweeds placed at 3 m inside and outside the edge of plots consisting of
placed at 1, 3, 5, and 10 m inside (negative value) and outside the field edge
Bt11, non-Bt (untreated), and non-Bt (-cyhalothrin-treated) sweet corn in
of event-176 field corn in Maryland. Means Ϯ SE are based on separate
Maryland. Data are based on separate bioassays conducted at 3, 6, and 9 days
bioassays conducted on days 3, 6, 9, and 14 of anthesis. Within each graph,
of anthesis. Within each graph, columns with the same lowercase letters are
columns with the same letters are not significantly different (P Ͻ 0.05; Tukey’s
not significantly different for the interaction effect; pairs of columns with the
same uppercase letters are not significantly different for the treatment effect(P Ͻ 0.05; Tukey’s test).
average of 0.9 mg, gained 9.9 to 12.8 mg after 4 days (Fig. 2). On
day 6, the plant position had a significant effect on larval survival
rately for each bioassay date and are summarized in Fig. 3. On
(P Ͻ 0.0001) and weight gain (P ϭ 0.0081). Sixty-three percent
all dates, the treatment by position-interaction effect on per-
of the larvae survived and gained 8.3 mg each after feeding on
centage of survival was significant (day 3, P Ͻ 0.0001; day 6, P Ͻ
leaves outside the field, compared with 25.1% survival and
0.0001; day 9, P ϭ 0.0027). Survival of monarch larvae feeding
weight gains of 4.8 mg inside. On day 9, larval survival inside the
on milkweed leaves either inside or outside Bt and non-Bt
field was reduced by 51% compared with survival of milkweeds
(unsprayed) plots ranged from 79.9 to 93.2% and were not
located outside (P ϭ 0.0215). Larvae also gained less weight
statistically different at any date. Survival of larvae feeding on
inside the field, but differences across plant positions were not
leaves collected inside and outside non-Bt (sprayed) plots was
significant. On day 14, weight gain, but not survival, was signif-
significantly reduced by 91–100% and 21–45%, respectively.
icantly affected by plant position (P ϭ 0.0418); however, differ-
Most larvae died within 1 h of exposure to sprayed leaves. Initial
ences across plant positions were not consistent with levels of
weights during days 3, 6, and 9 averaged 2.2, 2.2, and 3.5 mg per
pollen. This last bioassay used a combination of late-first and
larva, respectively. On day 3, the weight gain of surviving larvae
early-second instars with an initial weight of 5.6 mg per larva and
was significantly influenced by the treatment by position inter-
leaves from only four transects of plants. Thus, weight gains were
action (P ϭ 0.0003). Larvae fed milkweed leaves collected from
significantly higher compared with previous bioassays and ex-
inside unsprayed non-Bt plots exhibited the greatest weight gain,
whereas those larvae exposed to -cyhalothrin on leaves inside
Sweet-corn experiment. Maximum levels of pollen from both Bt
sprayed plots did not survive and, thus, showed no weight gain.
and non-Bt hybrids were deposited on milkweeds at 6 to 9 days
For bioassays conducted on days 6 and 9 of anthesis, the
after the onset of anthesis. Peak levels of pollen on milkweeds
treatment by position interaction and main effect for position
situated 3 m inside plots averaged 504–586 grains per cm2 and
were not significant. On day 6, only the treatment effect was
were significantly higher than levels on plants outside plots that
significant (P ϭ 0.0045); average weight gains of larvae feeding
peaked at 18–22 grains per cm2 (P Ͻ 0.0001). The overall level
on milkweeds from Bt plots were significantly higher than those
of pollen produced by both hybrids over time was statistically the
of larvae feeding on plants from non-Bt plots. The insecticide
same. Rain events, recorded on day 5 (2 mm) and day 9 (36 mm)
significantly reduced the weight gains of surviving larvae feeding
of anthesis, may have removed some pollen.
on plants from both inside and outside the sprayed non-Bt plots.
Data on larval survival and weight gain were analyzed sepa-
On day 9, overall weight gains of larvae after 4 days of feeding
11934 ͉ www.pnas.org͞cgi͞doi͞10.1073͞pnas.211277798
typically less diverse in flora and fauna than field edges, and thus
larvae may experience less predation within cornfields (13).
DiscussionThese studies provide evidence that the amount of pollen
deposited on milkweed leaves and the Cry1Ab expression in
pollen largely predict impacts of pollen on monarch larvae
feeding on milkweed associated with Bt corn during anthesis. In
the Iowa I study, low doses of event-176 pollen averaging 23
grains per cm2 had a significant effect on weights of first instars,
although survival differences were not statistically significant.
Results from the field-corn experiment conducted in Maryland
show that the survival and growth of first instars were signifi-
cantly affected when event-176 pollen levels reached Ϸ67 grains
per cm2 of leaf area. Survival of larvae feeding on milkweeds
within the field during peak anthesis was reduced by as much as
60%, and weight gain was also reduced by as much as 42%
compared with larvae feeding on milkweeds outside of the field.
These results are consistent with those of Jesse and Obrycki (5),
who observed a decrease in the survival of larvae exposed to
event-176 pollen on leaves from within a field (80–217 pollen
grains per cm2) compared with those fed leaves from outside of
the field. The results are also consistent with laboratory studies
that have shown reductions in weight gain at doses as low as 5–10
grains per cm2 after a 4-day exposure period (8).
In contrast, Mon810 and Bt11 hybrids do not seem to have
direct adverse effects on larval survival. The Iowa I study
demonstrates that low doses of Mon810 pollen (26 grains per
cm2) do not affect survivorship or weight gain of first instars after
a 5-day exposure period. The New York study also shows that
survival of developing larvae on milkweeds located within the
field was not significantly affected after exposure to Mon810
Survival curves for monarch larvae placed in and near Bt and non-Bt
pollen for 22 days. These results are not surprising, because
corn fields. (a) Iowa. (b) New York.
Mon810 pollen expresses far less toxin than event-176 pollen.
None of the studies show an effect of Bt11 pollen on survivorship
or weight gain either outside or inside the field, even where
were higher than the previous bioassays because of higher initial
pollen densities as high as 586 grains per cm2 of leaf area were
weights. A significant treatment effect (P ϭ 0.0148) indicated
observed. In the Ontario study, neither first nor third instars
that weight gains for the Bt and non-Bt (unsprayed) treatments
were detrimentally affected by pollen from event-Bt11 hybrids
were the same but significantly higher than average weights of
after exposure to pollen densities of 55 and 97 grains per cm2,
larvae feeding on milkweed leaves from both inside and outside
respectively. The survival, growth, and development of later-life
stages were also unaffected at these doses; larvae caged in Bt11
cornfields, non-Bt cornfields, or control areas developed into
Iowa II and New York Studies. Pollen densities among the three
pupae and adults of similar weight and size within similar
sites in Iowa were similar between hybrids and ranged from 154
developmental times. However, the percent of emergence may
to 367, 11 to 116, and 5 to 36 grains per cm2 within, at the edge,
have been affected by Bt11 pollen and warrants further inves-
and 2 m outside the field, respectively. There were no significant
tigation. In the Iowa II study, no effects on survival were
differences in the numbers of larvae surviving and in the survival
observed for larval cohorts feeding for 14 days on milkweed
curves among the three field sites. The number of larvae alive
plants within Bt11 corn plots with average pollen levels of 241
over the 14 days in the Bt and non-Bt corn plots was statistically
grains per cm2. Results from the sweet-corn experiment with the
the same. The survival curves of larvae pooled over the three Bt
Bt11 event show no effects on monarch survival or growth after
corn sites were not significantly different from those in non-Bt
4 days at the three bioassay times during anthesis, compared with
(Fig. 4a). In New York, trends in survivorship were also statis-
larvae in and near non-Bt plots that were not treated with
tically the same for cohorts of larvae feeding for 22 days on
insecticide. These field studies are consistent with those of
milkweeds in Bt and non-Bt fields (Fig. 4b). Pollen counts on
laboratory studies that suggest that exposure to Bt11 pollen at
milkweeds in the field averaged 127 grains per cm2 and were not
doses of less than 1,000 grains per cm2 do not detrimentally affect
significantly different between hybrids. The early-instar larvae
first-instar monarchs after a 4-day exposure period (8).
Most of the studies reported herein did not examine expo-
may have experienced higher pollen levels because counts were
sure in the middle of cornfields where pollen densities may be
made 6 days after larvae were introduced and Ͻ24 h after
higher than at the plot edge (14). However, in the sweet-corn
significant precipitation that may have reduced pollen levels
experiment, unaffected larvae fed on milkweed leaves with Ͼ3
(12). For data pooled over hybrids, the survivorship curves in
times the pollen densities typically found in field-corn plots. Bt
both studies indicated significantly lower survival on milkweeds
sweet corn can be viewed as a worst-case scenario corn type for
at the edge of the field compared with survival at the other
testing nontarget effects because it produces more pollen per
locations (Iowa: log-rank test, P Ͻ 0.09; Wilcoxon test, P Ͻ 0.04;
plant than field corn and is heavily treated with insecticides
New York: log-rank test, P Ͻ 0.009; Wilcoxon test, P Ͻ 0.04).
(http:͞͞www.epa.gov͞scipoly͞sap͞2000͞october͞). Further-
The same result has been observed in other studies for non-Bt
more, Cry1Ab expression in the pollen of Bt sweet corn is
fields and is probably explained by the fact that cornfields are
comparable to the expression in field-corn hybrids based on
PNAS ͉ October 9, 2001 ͉ vol. 98 ͉ no. 21 ͉ 11935
the Bt11 event. Thus, results of nontarget effects in sweet corn
recorded the fate of experimental cohorts of larvae for 14 to 22
can be extrapolated to risk scenarios for field corn.
days and show no apparent effect of Bt11 or Mon810 pollen on
The sweet-corn experiment also demonstrates the importance
survival, although the presence of other mortality factors con-
of assessing the risks of Bt corn to monarch populations in terms
tributed to high levels of variability in mortality rates; thus, subtle
of the relative risks of other agricultural practices. Monarch
effects of prolonged exposure to Bt toxin cannot be determined.
larvae were adversely affected by treatments of -cyhalothrin
Further, potential sublethal effects in response to long-term
applied to non-Bt plots of sweet corn. Most larvae died within
exposure, exposure of neonate larvae, and the potential impact
hours after feeding on milkweed leaves collected from plants
of Bt pollen on reproductive fitness and migration abilities could
exposed within plots to the spray application. Survival and
not be determined in these studies. Finally, because the detection
growth of larvae feeding on milkweeds outside of the sprayed
of very subtle effects is difficult in field studies because of the
plots also was reduced because of insecticide drift. These results
relatively small sample sizes, the results of these studies must be
were not surprising because -cyhalothrin is very effective at
considered with the results of much higher doses and more
controlling lepidopteran pests, and many reports have docu-
rigorous laboratory studies (8). In addition, the implications of
mented nontarget effects of conventional insecticides (14).
these studies must be understood in the context of the environ-
Recent data suggest that the use of Bt hybrids has the potential
mental doses anticipated throughout pollen dispersal (12) and
to significantly reduce the number of insecticide treatments
the likelihood that monarch larvae will be exposed to toxic doses
that are typically applied to sweet corn (http:͞͞www.epa.
gov͞scipoly͞sap͞2000͞october͞).
Differences in overall results of the Iowa I, Ontario, and
We thank Laura Timms, Pat Beaupre, Matt Van Ast, Bryan Muscat,
Maryland bioassays are clearly attributable to the protein ex-
Chad Harvey, Eric Olson, Terry Patton, Jeff Miner, Jessica Nelson,
pression level in pollen that is regulated by the transformation
Keith Bidne, Randy Ritland, Jim Robbins, Colothdian Tate, Patricia
event. Corn hybrids based on event 176 may be hazardous to
Anderson, Denny Bruck, Stacy van Loon, Kate Kronback, Jaleen
susceptible stages of monarch larvae that are present on their
Bruner, Karen Douchette, Kerry Gillooly, Erin Roe, Tegwin Taylor,
host plants within cornfields during anthesis. However, the
Maureen Carter, Lee Macomber, Jeremiah Depue, Christine Cappa-
dora, Jeffrey Fuchsberg, and Christa Hoffman for their assistance in
exposure dose of Bt11 pollen present at peak anthesis and the
conducting field trials. We also thank Fred Gould, George Kennedy,
Cry1Ab concentration in pollen ingested over the 4- or 5-day
Kevin Steffey, Anthony Shelton, Jeffrey Wolt, and Eric Sachs for their
feeding period was not high enough to significantly affect larval
critical reviews and Orley Taylor (Monarch Watch, University of Kansas)
survival. These findings are probably true for Mon810 as well,
for providing monarch larvae. This research was supported by a pooled
although it was not vigorously tested, because Cry1Ab expres-
grant provided by the United States Department of Agriculture, Agri-
sion in Mon810 is similar to that in Bt11 pollen. However, further
cultural Research Service, and the Agricultural Biotechnology Steward-
research is required to better understand the impact of Bt pollen
ship Technical Committee (ABSTC), and by funding from the Canadian
with relatively low toxicity on monarch populations. For exam-
Food Inspection Agency, Environment Canada, the Ontario Corn
ple, in the natural setting, larvae hatching at the onset of anthesis
Growers’ Association, the Maryland Agricultural Experiment Station,
and the Leopold Center for Sustainable Agriculture (Ames, IA). Mem-
may be exposed to biologically active Cry1Ab in pollen for a
bers of ABSTC are Aventis CropScience USA LP, Dow AgroSciences
longer period than 4 or 5 days; thus, exposure duration needs to
LLC, E. I. du Pont de Nemours and Company, Monsanto Company, and
be considered along with exposure concentration as a determi-
Syngenta Seeds, Inc. L.C.H.J. was supported by an Environmental
nant of environmental dose. The Iowa II and New York studies
1. Koziel, M. G., Beland, G. L., Bowman, C., Carozzi, N. B., Crenshaw, R.,
9. Raynor, G. S., Ogden, E. C. & Hayes, J. V. (1972) Agron. J. 64,
Crossland, L., Dawson, J., Desai, N., Hill, M., Kadwell, S., et al. (1993)
Biotechnol. 11, 194–200.
10. Russek-Cohen, E. & Douglas, L. W. (1999) Mixed Model Short Course Manual
2. Schuler, T. H., Poppy, G. M., Kerry, B. R. & Denholm, I. (1999) Trends
(University of Maryland, Beltsville), pp. 89–103. Biotechnol. 17, 210–216.
11. SAS Institute (1990) SAS͞STAT User’s Guide, Version 6.0 (SAS Inst., Cary,
3. Fearing, P. L., Brown, D., Vlachos, D., Meghji, M. & Privalle, L. (1997) Mol.Breeding 3, 169–176.
12. Pleasants, J. M., Hellmich, R. L., Dively, G., Sears, M. K., Stanley-Horn, D. E.,
4. Losey, J. E., Rayor, L. S. & Carter, M. E. (1999) Nature (London) 399,214.
Mattila, H. R., Foster, J. E., Clark, P. L. & Jones, G. D. (2001) Proc. Natl. Acad.
5. Jesse, L. C. H. &. Obrycki, J. J. (2000) Oecologia 125, 241–248. Sci. USA 98, 11919–11924. (First Published September 14, 2001; 10.1073͞
6. Malcolm, S. B. & Brower, L. P. (1989) Experientia 45, 284–294.
7. Wassenaar, L. I. & Hobson, K. A. (1998) Proc. Natl. Acad. Sci. USA 95,
13. Oberhauser, K. S., Prysby, M., Mattila, H. R., Stanley-Horn, D. E., Sears, M. K.,
Dively, G., Olson, E., Pleasants, J. M., Lam, W.-K. F. & Hellmich, R. L. (2001)
8. Hellmich, R. L., Siegfried, B., Sears, M. K., Stanley-Horn, D. E., Daniels, M. J.,
Proc. Natl. Acad. Sci. USA 98, 11913–11918. (First Published September 14,
Mattila, H. R., Spencer, T., Bidne, K. G. & Lewis, L. (2001) Proc. Natl. Acad. Sci.USA 98, 11925–11930. (First Published September 14, 2001; 10.1073͞
14. Jepson, P. C. (1989) Pesticides and Non-Target Invertebrates (Intercept, An-
11936 ͉ www.pnas.org͞cgi͞doi͞10.1073͞pnas.211277798
Medical Information Database Name: __________________________________________ Date: __________________ Reason for seeing doctor: ___________________________________________________ Referring Physician: _______________________________________________________ Were you seen in the Emergency Room? YES NO ___ Richland ___ Baptist ___ Lexington ___ Providence ___ Providence NE ___ Other Date
“Vitamin D is like a Religious Revival; either you're a believer or Adam Thornton discusses vitamin D with Dr Michael Holick There is no doubt that the current ‘vitamin du jour’ in both the scientific literature and the lay media is vitamin D. Vitamin D is rightly in the spotlight because of the enormous amount of scientific research that has been conducted in recent years that sheds light
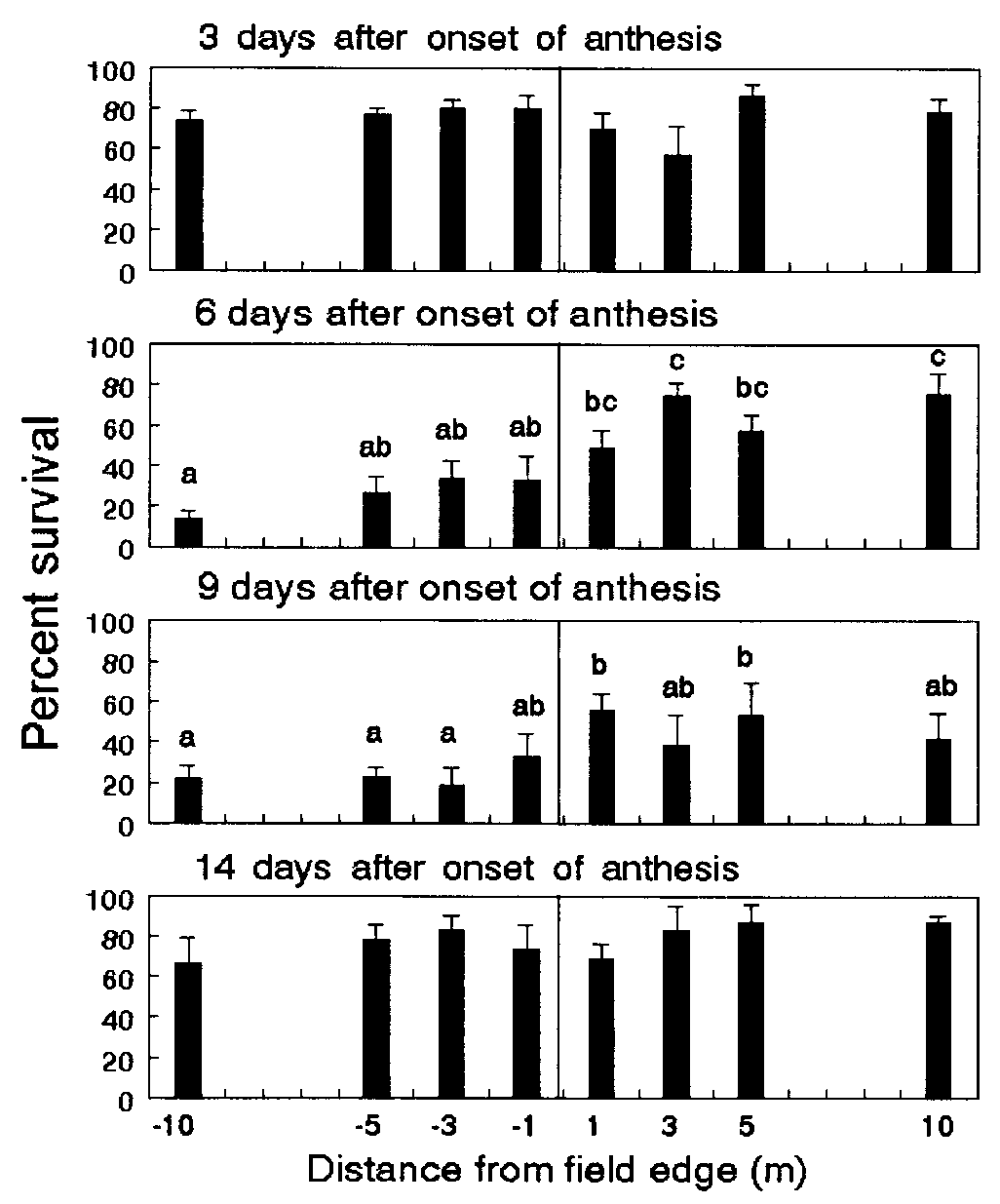 Table 1. Comparison of survivorship and growth for Monarch larvae exposed as first or third instars to Bt11 and non-Bt pollen on
Table 1. Comparison of survivorship and growth for Monarch larvae exposed as first or third instars to Bt11 and non-Bt pollen on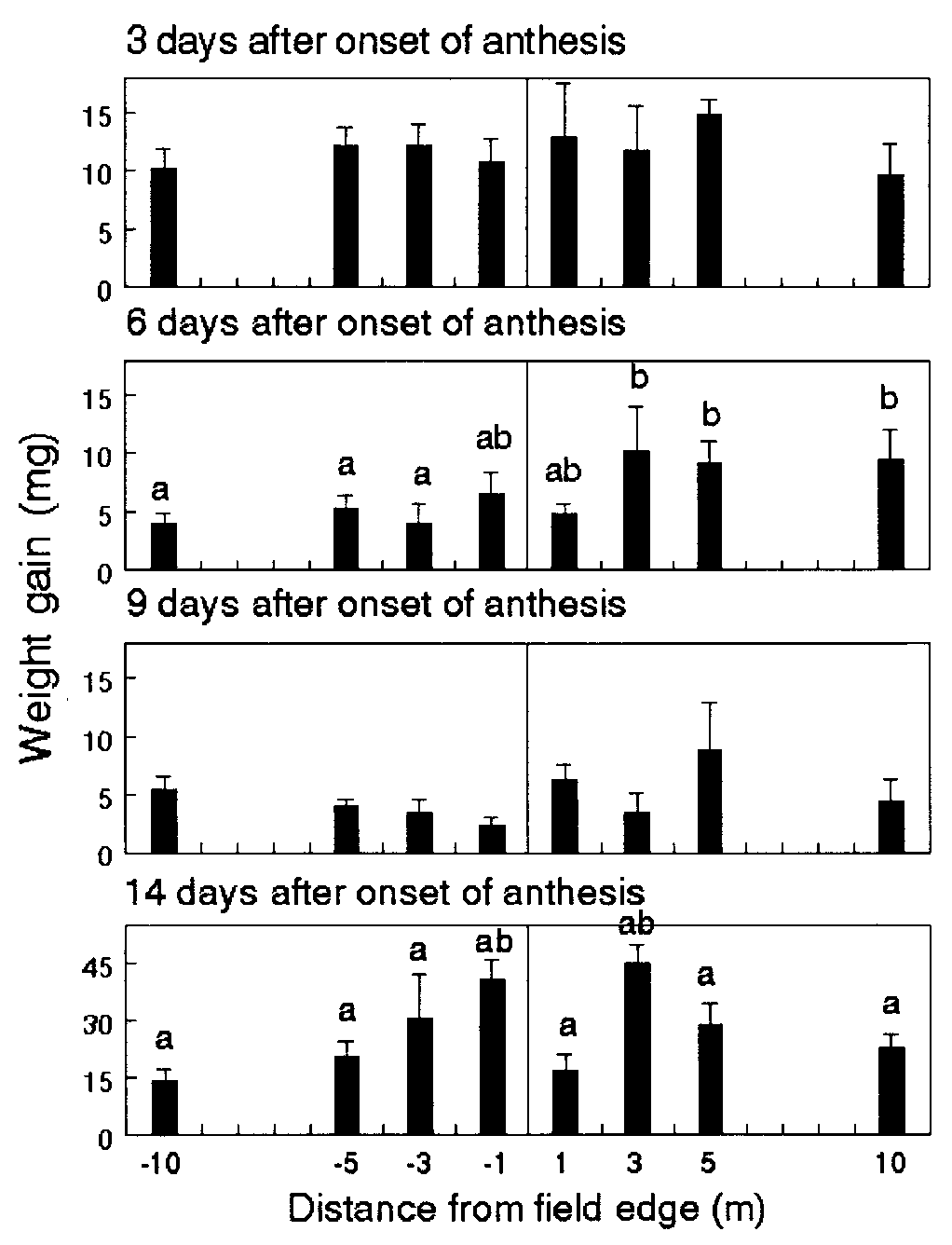
 Survival and weight gain of first-instar monarch larvae feeding on
Weight gain of first-instar monarch larvae feeding on milkweeds
milkweeds placed at 3 m inside and outside the edge of plots consisting of
placed at 1, 3, 5, and 10 m inside (negative value) and outside the field edge
Bt11, non-Bt (untreated), and non-Bt (-cyhalothrin-treated) sweet corn in
of event-176 field corn in Maryland. Means Ϯ SE are based on separate
Maryland. Data are based on separate bioassays conducted at 3, 6, and 9 days
bioassays conducted on days 3, 6, 9, and 14 of anthesis. Within each graph,
of anthesis. Within each graph, columns with the same lowercase letters are
columns with the same letters are not significantly different (P Ͻ 0.05; Tukey’s
not significantly different for the interaction effect; pairs of columns with the
same uppercase letters are not significantly different for the treatment effect(P Ͻ 0.05; Tukey’s test).
Survival and weight gain of first-instar monarch larvae feeding on
Weight gain of first-instar monarch larvae feeding on milkweeds
milkweeds placed at 3 m inside and outside the edge of plots consisting of
placed at 1, 3, 5, and 10 m inside (negative value) and outside the field edge
Bt11, non-Bt (untreated), and non-Bt (-cyhalothrin-treated) sweet corn in
of event-176 field corn in Maryland. Means Ϯ SE are based on separate
Maryland. Data are based on separate bioassays conducted at 3, 6, and 9 days
bioassays conducted on days 3, 6, 9, and 14 of anthesis. Within each graph,
of anthesis. Within each graph, columns with the same lowercase letters are
columns with the same letters are not significantly different (P Ͻ 0.05; Tukey’s
not significantly different for the interaction effect; pairs of columns with the
same uppercase letters are not significantly different for the treatment effect(P Ͻ 0.05; Tukey’s test).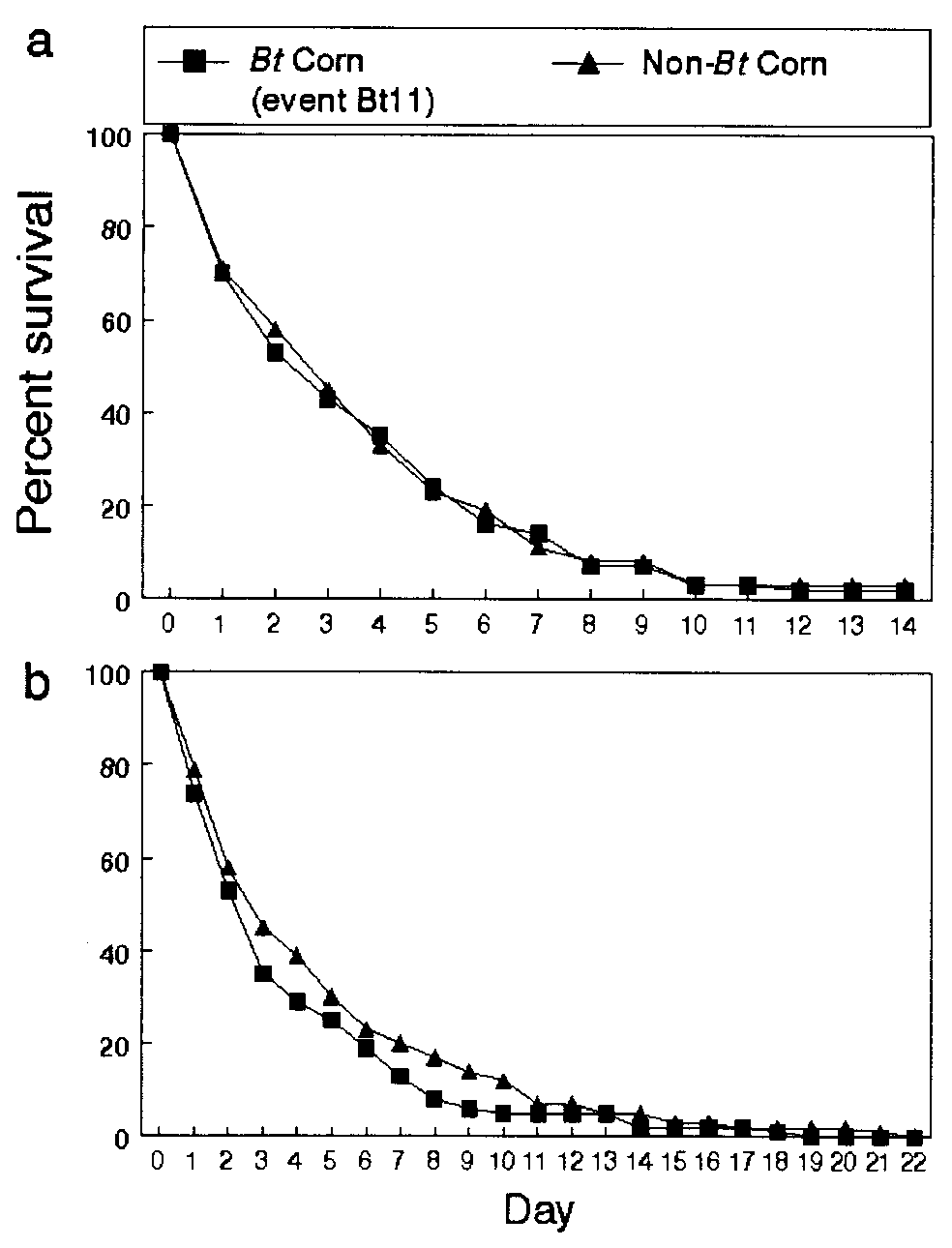 typically less diverse in flora and fauna than field edges, and thus
larvae may experience less predation within cornfields (13).
typically less diverse in flora and fauna than field edges, and thus
larvae may experience less predation within cornfields (13).