Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Norgestomet and estradiol valerate induced luteolysis is dependent upon the uterus
NORGESTOMET AND ESTRADIOL VALERATE INDUCED LUTEOLYSIS IS
C. A. Peterson, J. C. Huhn, and D. J. Kesler
SUMMARY Beef heifers were assigned to three groups: 1) untreated controls (n= 4), 2) Syncro-Mate B® (SMB) treated (n= 5), and 3) hysterectomized and SMB treated (n= 4). SMB was administered eight or nine days after estrus, approximately 30 days after hysterectomy. This study was conducted to determine if the uterus was necessary for SMB to induce luteolysis. SMB induced premature luteolysis as only 20% of the intact SMB treated heifers had ≥ .75 ng/mL of progesterone seven days after the time of SMB treatment compared to all (100%) of the untreated heifers (P <.05). By nine days after the time of SMB treatment, 25% of the untreated heifers and none (0%) of the intact SMB treated heifers had ≥ .75 ng/mL of progesterone; however, all (100%) of the hysterectomized SMB treated heifers had ≥ .75 ng/mL of progesterone (P <.05). Therefore, SMB-induced luteolysis required the involvement of the uterus. The luteolysin, prostaglandin F2α, is probably the secretion from the uterus that mediates the SMB-induced luteolysis. SMB treatment, however, required 7-8 days to induce luteolysis. INTRODUCTION Syncro-Mate B® (SMB) is a commercially available procedure to synchronize estrus in beef and dairy cattle. The procedure consists of a norgestomet implant and an intramuscular injection containing norgestomet and estradiol valerate administered at the time of implantation. SMB has three known mechanisms of action. First, an estrus suppression dosage of norgestomet diffuses from the implant during the nine days in situ (Kesler and Favero, 1995). Secondly, the injection causes atresia of antral follicles and recruitment of a new cohort of follicles four to five days after administration (Vasconcelos et al., 1997). Thirdly, the injection causes regression of corpora lutea (Kesler and Favero, 1995). Since the implant is left in place for nine days, the injection is needed to induce regression of corpora lutea in cows during the first half of the estrous cycle. Estradiol-17β, the active metabolite of the estradiol valerate contained within the SMB injection, has been demonstrated to hasten corpus luteum regression (Thatcher et al., 1986). Thatcher et al. (1986) reported spikes of 15-keto- 13, 14-dihydro-prostaglandin F2α (PGFM) in the peripheral blood before luteolysis ensued and concluded that estradiol-17β induced luteolysis by provoking a release of PGF2α from the uterus; however, Thatcher et al. (1986) administered estradiol-17β during the second half of the estrous cycle. Progesterone treatment during metestrus has also been reported to shorten the estrous cycle, but only by four days (Woody et al., 1967; Harms and Malven, 1969; Ginther, 1970; Battista et al., 1984; Garrett et al., 1988). The objective of this study was to determine if the hypothesis that SMB induced luteolysis is dependent upon uterine involvement was correct. MATERIALS AND METHODS Three groups of purebred Angus beef heifers from the University of Illinois beef research unit (Urbana, IL) were included in this study. The control group (n=4) was selected from a larger group of estrus-cycling females administered prostaglandin F2α (25 mg Lutalyse®; Pharmica and Upjohn, Kalamazoo, MI, USA) due to their similar timing of estrus (detectable estrus within 48 hours of each
other). They were observed for estrus twice daily, at least 30 minutes in the morning, and again in the evening. Estrus was defined as a female standing to be mounted by another animal. The second group of females (n=5) that was similarly selected from a larger group of estrus-cycling females, were treated with SMB (Merial, Inc., Athens, GA, USA) on day eight or nine of the estrous-cycle. SMB treatment included a hydron ear implant that contained 6.0 mg norgestomet, along with an intramuscular injection of norgestomet (3.0 mg) and estradiol valerate (5.0 mg) in sesame seed oil and benzyl alcohol. The ear implant was administered subcutaneously in the convex surface of the middle one-third of the ear. Nine days after implantation, the implants were removed. The third group consisted of four heifers. They were hysterectomized leaving the ovaries and ovarian vasculature intact. After preparation for surgery, heifers were placed in dorsal recumbency to allow access to the surgical site. The incision was made from the umbilicus to the cranial aspect of the pubis. A self-retaining abdominal retractor was used to facilitate exposure of the uterus. The uterus was then mobilized from the normal in situ position and retracted through the abdominal incision. It was then ligated immediately cranial to the internal os of the cervix with a surgical stapling instrument. The broad ligament of the uterus was incised cranially until the utero-ovarian ligament was located. Once the ovary and its blood supply were isolated, using a crushing hemostatic forcep, the utero-ovarian ligament was detached. The same procedure was preformed on both uterine horns. The uterus was then removed from the abdomen, and the abdominal area was then closed in four layers. After approximately 20 days of recovery, the hysterectomized females were intramuscularly administered a luteolytic dose of PGF2α (25 mg Lutalyse®; Pharmica and Upjohn, Kalamazoo, MI). The females were observed for estrus and SMB was administered on days eight or nine of the estrous cycle, following the same procedures for SMB as described earlier. All hysterectomized heifers exhibited estrus during the same 48 hour period as described for the untreated and SMB treated heifers. Heifers in all three groups were bled by jugular venipuncture with 3.75 cm, 18 g needles and 10 cc syringes. The control and the SMB treated heifers were bled daily, from day eight or nine through day 17 or 18 of the estrous cycle, while the hysterectomized heifers were bled daily from day eight or nine through day 19 or 20 of the estrous cycle. Within two hours of collection, all blood samples were centrifuged at 2,000 x g and stored at –20° C until assayed. All samples were assayed for progesterone concentrations by a validated enzyme immunoassay (Kesler et al., 1990) and data were analyzed by split-plot analysis of variance as described by Gill and Hafs (1971). The percent of heifers that experienced luteolysis, a decrease in progesterone concentrations to < .75 ng/mL, was analyzed by chi square analysis as described by Cochran and Cox (1957). RESULTS Luteolysis, a decrease in progesterone concentrations < .75 ng/mL, did not occur in any of the untreated heifers until nine days after the time of SMB treatment, day 17-18 of the estrous cycle (Table 1). Luteolysis occurred in one, four, and five of the five intact SMB treated heifers, by days 6, 7, and eight, respectively. SMB did not induce luteolysis in any of the hysterectomized heifers; therefore, SMB induced a lower (P<.05) incidence of luteolysis in hysterectomized heifers than in intact heifers. As illustrated in Figure 1, progesterone concentrations were sustained in the hysterectomized heifers administered SMB, whereas progesterone concentrations fell at seven days post-SMB treatment in the intact heifers administered SMB (group P<.05; group x time interaction P<.01).
DISCUSSION During the nine days in situ, the SMB implant releases a dose, ≥140 µg per day, of norgestomet that effectively suppresses estrus (Machado and Kesler, 1996). Because implants are left in situ only for nine days, females treated during the first half of the estrous cycle also require a compound that causes premature corpus luteum regression. The norgestomet released from the SMB implant has no deleterious effects on corpora lutea formation and function even when administered during metestrus (Kesler, 1997; Kesler et al., 1997). The injection administered at the time of insertion of the SMB implant contains 5.0 mg of estradiol valerate and 3.0 mg of norgestomet. Data demonstrate that SMB causes premature corpus luteum regression (Kesler and Favero, 1995; Kesler and Favero, 1996); however, the mechanism by which SMB induces corpus luteum regression has not been elucidated. Since administration of the implant alone has no deleterious effects on corpus luteum formation and function, we conclude that it is the injection that causes premature corpus luteum regression. Lemon (1975) suggested that the estradiol valerate and norgestomet injection may cause corpus luteum regression through negative feedback on luteinizing hormone (LH) release-an anti-luteotropic effect. It is doubtful that this hypothesis is correct. Burns et al. (1993) demonstrated that although LH was immediately inhibited, this decrease in LH did not totally abolish corpus luteum development. Furthermore, Peters et al. (1994) demonstrated that abolishing the pulse releases of LH with a gonadotropin releasing hormone antagonist did not affect corpus luteum lifspan. An alternative hypothesis is that the SMB injection causes luteolysis and this hypothesis has supporting data. Thatcher et al. (1986) administered estradiol-17β (3 mg) and hastened corpus luteum regression. In that study, corpus luteum regression, which occurred about 97 hours after estradiol-17β treatment, was proceeded by spikes of PGFM, the primary stable metabolite of PGF2α (Thatcher et al., 1986). Estradiol valerate, the estrogen in the SMB injection, is an estradiol-17β ester. Estradiol valerate has prolonged activity and elevates blood estradiol concentrations for seven to 12 days (Sinkula, 1978; Ashimine et al., 1991); however, it is converted to estradiol-17β before it is biologically active (Kesler et al., 1980). Exogenous progesterone administration during metestrus has also been demonstrated to reduce the interestrous interval by four days (Woody et al., 1967; Harms and Malven, 1969; Ginther, 1970; Battista et al., 1984; Garrett et al., 1988). It has been suggested that the decreased interestrous interval after treatment with exogenous progesterone is mediated through earlier maturation of the uterus as indicated by an earlier release of PGF2α from the uterine endometrium (Garrett et al., 1988). Wiltbank and Casida (1957) demonstrated that the lifespan of corpora lutea could be extended by hysterectomy. Furthermore, Warren et al. (1979) demonstrated that the lifespan of corpora lutea could be extended by indomethacin, a biosynthetic inhibitor to PGF2α synthesis, infusion into the uterine horn ipsilateral to the ovary with the corpus luteum. Collectively, the results of this study with the results of others, as previously described, suggest that the SMB injection is probably causing the uterine endometrium to release PGF2α prematurely. This premature release of PGF2α causes corpus luteum lysis during the SMB implantation period so that at the end of the nine-day implantation period, all females are relying on the norgestomet from the SMB implant to suppress estrus. The delayed regression in corpora lutea in SMB treated heifers as
compared with estradiol-17β treated heifers (approximately four days vs. approximately seven days, respectively) may be due to delayed availability of estradiol-17β from the estradiol valerate injection and/or inclusion of a progestin (Garrett et al., 1988). However, a delayed response is necessary as females treated on days one or two of the estrous cycle require two to three days of additional development of the corpus luteum before PGF2α can provoke luteolysis. Available data suggest that all cattle, except metestrus cattle, have a high estrus synchronization rate to SMB treatment (Kesler and Favero, 1996). Metestrus females, however, only have an approximately 50% estrus synchronization response rate to SMB (Kesler and Favero, 1995). In metestrus females not responding to SMB, the injection does not induce corpus luteum regression. Therefore, the norgestomet and estradiol valerate injection is less effective in inducing corpus luteum regression in metestrus females; however, inclusion of a larger quantity of norgestomet appears to improve the efficacy of corpus luteum lysis in metestrus females (Fanning et al., 1992). Although norgestomet is far more biologically active than progesterone (Gilbert et al., 1974), it is metabolized quickly (Chien, 1978). In the studies that were conducted to determine that progesterone caused an earlier release of PGF2α from the uterine endometrium and reduced the interestrous interval, progesterone was administered daily for four days (Garrett et al., 1988). In summary, SMB-induced luteolysis appears to be due to the norgestomet and estradiol valerate from the SMB injection and requires involvement of the uterus. We suggest that the injection causes a delayed but permature release of PGF2α from the uterine endometrium. LITERATURED CITED Ashimine, D., Miller-Patrick, K., Webster, H., Weems, C, Vincent D, 1991. Sources of increased
estradiol (E) following Syncro-Mate B (SMB) in beef heifers. J. Anim. Sci. 69 (Suppl 1), 400.
Battista, P.J., Rexroad, C.E., Williams, W.F., 1984. Effects of progesterone administered to dairy
heifers on sensitivity of corpora luteal to PGF2α and on plasma LH concentration. Theriogenology 22, 47.
Burns, P.D., Spitzer, J.C., Bridges, Jr., W.C., Hendricks, D.M., Pyler, B.B., 1993. Effects of
metestrus administration of a norgestomet implant and injection of norgestomet and estradiol valerate on luteinizing hormone release and development and function of corpora lutea in suckled beef cows. J. Anim. Sci. 71, 983.
Chien, Y.E., 1978. Methods to achieve sustained drug delivery. A physical approach: implants. In:
Sustained and Controlled Release Drug Delivery Systems. Ed., J.R. Robinson. Marcel Dekker, Inc., New York, p. 328.
Cochran, W.G., Cox, G.M., 1957. Experimental designs, 2nd ed., John Wiley and Sons., Inc., New
Fanning, M.D., Spitzer, J.C., Burns, G.L., Plyler, B.B., 1992. Luteal function and reproductive
response in suckled beef cows after metestrus administration of a norgestomet implant and
injection of estradiol valerate with various dosages of injectable norgestomet. J. Anim. Sci. 70, 1352.
Garrett, J.E., Geisert, R.D., Zavy, M.T., Gries, L.K., 1988. Effect of exogenous progesterone on
prostaglandin F2α release and the interestrous interval in the bovine. Prostaglandins 36, 85.
Gilbert, H.G., Phillips, G.H., English, A.F., 1974. The progestational and anti-estrogenic activities
of some novel 11β-substituted steroids. Steroids 23, 585.
Gill, J.L., Hafs, H.D., 1971. Analysis of repeated measurements of animals. J. Anim. Sci. 33, 331. Ginther, O.J., 1970. Effect of progesterone on length of estrous cycle in cattle. Amer. J. Vet. Res.
Harms, P.G., Malven, P.V., 1969. Modification of the bovine luteal function by exogenous oxytocin
Kesler, Darrel J. 1997. Remotely-delivered contraception with needle-less norgestomet implants.
In: Contraception in Wildlife Management, Ed., T.J. Kreeger, U.S. Government Printing Office, Washington, D.C., pp. 171-184.
Kesler, D.J., Faulkner, D.B., Nash, T.G., 1997. Norgestomet implantation in beef females. Large
Kesler, D.J., Favero, R.J., 1995. Estrus synchronization in beef females with norgestomet and
estradiol valerate. Part 1: Mechanism of action. Agri-Practice 16, 10.
Kesler, D.J., Favero, R.J., 1996. Estrus synchronization in beef females with norgestomet and
estradiol valerate. Part 2: Factors limiting and enhancing efficacy. Agri-Pract. 17, 12.
Kesler, D.J., Khazali, H. , Favero, R. J., 1990. Quantification of steroids via a polymer linked
second antibody immunoassay: methods of linking anti-rabbit IgG to poly(styrene). In: Progress in Biomedical Polymers. Ed., C. G. Gebelein and R. Dunn. Plenum Publishing Co., New York, p. 157.
Kesler, D.J., Troxel, T.R., Vincent, D.L., Carlin, S.E., 1980. Luteinizing hormone concentrations in
anestrous ewes administered various estrogens. Theriogenology 13, 231.
Lemon, M., 1975. The effect of oestrogens alone or in association with progestagens on the
formation and regression of the corpus luteum of the cyclic cow. Ann. Biol. Anim. Bioch. Biophys. 15, 243.
Machado, R., Kesler. D.J., 1996. Efficacy of norethindrone acetate and norgestomet implants in
suppressing estrus in female beef cows. Drug Dev. Ind. Pharm. 22, 1211.
Peters, K.E., Bergfeld, E.G., Cupp, A.S., Kojima, F.N., Mariscal, V., Sanchez, T., Wehrman, M.E.,
Grotjan, H.E., Hamernik, D.L., Kittok, R.J., Kinder, J.E., 1994. Luteinizing hormone has a role in development of fully functional corpora lutea (CL) but is not required to maintain CL function in heifers. Biol. Reprod. 51, 1248.
Sinkula, A.A., 1978. Methods to achieve sustained drug delivery. A chemical approach. In:
Sustained and Controlled Release Drug Delivery Systems. Ed., J.R. Robinson. Marcel Dekker, Inc., New York, p. 411.
Thatcher, W.W., Terqui, M., Thimonier, J., Mauleon , P., 1986. Effect of estradiol-17β on
peripheral plasma concentration of 15-keto-13,14-dihydro PGF2α and luteolysis in cyclic cattle. Prostaglandins 31, 745.
Vasconcelos, J.L.M., Silcox, RW., Pursley, J.R., Wiltbank M.C., 1997. Effect of day of estrous
cycle on synchronization of ovulation using GnRH. J. Anim. Sci. 75 (Suppl. 1), 89 (Abstr.).
Warren, J.E., Lewis, P.E., Kiesling, D.O., 1979. Inhibition by indomethacin of estrogen induced
luteal regression in heifers. J. Anim. Sci. Vol. 49 (Suppl.1), 346 (Abstr.).
Wiltbank, J.N., Casida, L.E., 1957. Alteration of ovarian activity by hysterectomy. J. Anim. Sci.
Woody, C.D., First, N.L., Pope, A.L., 1967. Effect of exogenous progesterone on estrous cycle
Figure 1. Progesterone concentrations of SMB treated heifers (dotted line) and hysterectomized/SMB treated heifers (solid line) administered SMB on days 8 or 9 of the estrous cycle.
Table 1. Number (percent) of untreated heifers, SMB-treated heifers, and SMB-treated hysterectomized heifers with progesterone concentrations ≥.75 ng/mL on days 6 through 9 of the estrous cycle. Days Post-Treatment-
Day 6 Post-Treatment- day 14-15 of the estrous cycle
Day 7 Post-Treatment- day 15-16 of the estrous cycle
Day 8 Post-Treatment- day 16-17 of the estrous cycle
Day 9 Post-Treatment- day 17-18 of the estrous cycle
x,yValues within the same row with different superscripts differ (P < .01).
y,zValues within the same row with different superscripts differ (P < .05).
Effects of calcium alkaline ionized water on formation and maintenance of osseous tissues (osteoporosis). Rei Takahashi, Zhenhua Zhang, Yoshinori ItokawaKyoto University Graduate School of Medicine, Dept. of Pathology and Tumor Biology, Fukui Prefectural University, Japan. "Effects of calcium alkaline ionized water on formation and maintenance of osseous tissues in rats were examined.
November 11, 1998 N.G.I.S.C. Las Vegas Meeting CHAIRPERSON JAMES: I want to thank each and everyone of our panelists. At this point we will open it up fordiscussion, any questions from our commissioners and even anexchange among yourselves if you would like to do that. DR. DOBSON: Doctor Nora, yesterday Mitzi Schlichtermade a passing reference to medication for her husband, Art whohas b
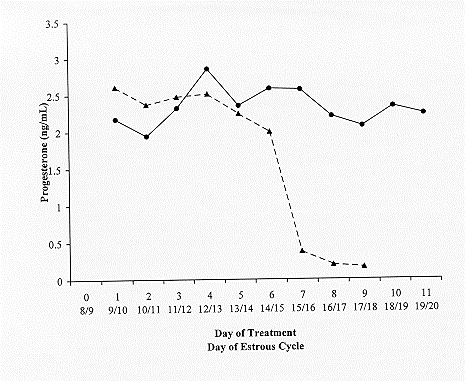 Figure 1. Progesterone concentrations of SMB treated heifers (dotted line) and hysterectomized/SMB treated heifers (solid line) administered SMB on days 8 or 9 of the estrous cycle.
Table 1. Number (percent) of untreated heifers, SMB-treated heifers, and SMB-treated hysterectomized heifers with progesterone concentrations ≥.75 ng/mL on days 6 through 9 of the estrous cycle. Days Post-Treatment-
Day 6 Post-Treatment- day 14-15 of the estrous cycle
Day 7 Post-Treatment- day 15-16 of the estrous cycle
Day 8 Post-Treatment- day 16-17 of the estrous cycle
Day 9 Post-Treatment- day 17-18 of the estrous cycle
x,yValues within the same row with different superscripts differ (P < .01).
y,zValues within the same row with different superscripts differ (P < .05).
Figure 1. Progesterone concentrations of SMB treated heifers (dotted line) and hysterectomized/SMB treated heifers (solid line) administered SMB on days 8 or 9 of the estrous cycle.
Table 1. Number (percent) of untreated heifers, SMB-treated heifers, and SMB-treated hysterectomized heifers with progesterone concentrations ≥.75 ng/mL on days 6 through 9 of the estrous cycle. Days Post-Treatment-
Day 6 Post-Treatment- day 14-15 of the estrous cycle
Day 7 Post-Treatment- day 15-16 of the estrous cycle
Day 8 Post-Treatment- day 16-17 of the estrous cycle
Day 9 Post-Treatment- day 17-18 of the estrous cycle
x,yValues within the same row with different superscripts differ (P < .01).
y,zValues within the same row with different superscripts differ (P < .05).