Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Pii: s0891-5849(01)00477-4
Free Radical Biology & Medicine, Vol. 30, No. 7, pp. 803– 808, 2001
Copyright 2001 Elsevier Science Inc. PII S0891-5849(01)00477-4 Fast Track Paper
NITROXYL OXIDIZES NADPH IN A SUPEROXIDE DISMUTASE
NDREAS REIF,* LUIGI ZECCA,*† PETER RIEDERER,* MARTIN FEELISCH,‡ and HARALD H. H. W. SCHMIDT
*Clinical Neurochemistry, Department of Psychiatry and Psychotherapy, Julius-Maximilians-University, Wu¨rzburg, Germany;
†Institute of Advanced Biomedical Technologies, National Research Council, Segrate, Italy; ‡Department of Molecular and
Cellular Physiology, Louisiana State University Health Sciences Center, Shreveport, LA, USA; and §Rudolf-Buchheim-Institute for
Pharmacology, University of Gießen, Gießen, Germany
(Received 14 July 2000; Revised 28 November 2000; Accepted 16 January 2001)
Abstract—Nitric oxide synthases (NOS) convert L-arginine and N-hydroxy-L-arginine to nitric oxide (•NO) and/or nitroxyl (NOϪ) in a NADPH-dependent fashion. Subsequently, •NO/superoxide (OϪ-derived peroxynitrite (ONOOϪ)
consumes one additional mol NADPH. The related stoichiometry of NOϪ and NADPH is unclear. We here describe thatNOϪ also oxidizes NADPH in a concentration-dependent manner. In the presence of superoxide dismutase (SOD),which also converts NOϪ to •NO, nitrite accumulation was almost doubled and no oxidation of NADPH was observed. Nitrate yield from NOϪ was low, arguing against intermediate ONOOϪ formation. Thus, biologically formed NOϪ mayfunction as an effective pro-oxidant unless scavenged by SOD and affect the apparent NADPH stoichiometry of theNOS reaction. Keywords—NO synthase, Nitroxyl, NADPH, Superoxide dismutase, Nitrite, Nitrate, Peroxynitrite, Free radicals INTRODUCTION
pal products which may be converted into •NO have beensuggested, including nitroxyl (NOϪ) [11,12]. As NOϪ
Nitrogen monoxide (nitric oxide, •NO) is a physiologi-
can be converted to •NO by stoichiometric amounts of
cally and pathophysiologically important messenger
SOD [12,13], principal NOϪ formation would explain
molecule synthesized by a family of NO synthases
why •NO can only be detected in the presence of SOD.
[NOS; L-arginine, NADPH:oxygen oxidoreductases (ni-
Alternatively, it was suggested that NOS produces su-
tric oxide forming); EC 1.14.23.39]. These multifunc-
peroxide [14,15], which rapidly reacts with •NO to form
tional enzymes require the presence of NADPH, FAD,
oxoperoxonitrate(1Ϫ) (peroxynitrite, ONOOϪ) [16]. Un-
FMN, heme, and tetrahydrobiopterin (H Bip) [1,2], a
der these conditions, SOD would be needed to scavenge
structural Zn, as well as the substrates L-arginine and
superoxide in order to enable the detection of •NO.
molecular oxygen [3–7]. NADPH donates electrons for
However, such a role of SOD would not explain why the
the reduction of FAD and FMN [8], which function as
absolute amounts of enzyme (5 kU mlϪ1) required to
prosthetic groups shuttling the electrons to the heme
detect NO from purified NOS are far in excess of the
where molecular oxygen is activated [9].
amounts required to catalyze the dismutation of super-
Numerous studies were carried out in order to mea-
sure directly •NO formation from NOS either electro-
To resolve this issue, it is important to determine the
chemically or by gas phase chemiluminescence [10 –12].
NADPH stoichiometry of the principal NOS reaction.
However, when purified neuronal NOS (NOS-I) was
Literature values differ between 1, 1.5, and 2 mol
used, the presence of supracatalytic amounts of SOD was
NADPH per mol L-citrulline [17–19], depending on the
required to detect •NO [10,12]. Thus, alternative princi-
presence of cofactors in the incubation media. We re-cently reported an NADPH/citrulline-stoichiometry of
Address correspondence to: Dr. Andreas Reif, Department of Psy-
1.6, which was significantly lowered to 0.8 by SOD [12].
chiatry and Psychotherapy, Julius-Maximilians-University, Fu¨chslein-
A NOS-derived N-oxide may interfere with the observed
str. 15, D-97080 Wu¨rzburg, Germany; Tel: ϩ49 (931) 201-7672; Fax:
ϩ49 (931) 201-7755; E-Mail: a.reif@gmx.net.
NADPH stoichiometry, as ONOOϪ was shown to con-
sume one mol NADPH [12]. Moreover, Marletta and
lowing modifications. Aliquots of the Angeli’s salt stock
coworkers recently examined the reactions of H Bip-free
solution (100, 200, or 400 M) were added to 0.5 M
NOS [20] utilizing N-hydroxy-L-arginine, the proposed
TEA buffer (100 l, pH 7.0). The samples were then
intermediate of the NOS-reaction [17], as a substrate.
incubated for 5 min at 37°C in the absence or presence of
Under these conditions, NOϪ was formed with a stoichi-
100 M NADPH, with or without 5 kU mlϪ1 SOD.
ometry of 1.5:1 NADPH/NOϪ, whereas the theoretical
Thereafter, samples were frozen in liquid nitrogen and
value for this reaction should be 1:1. Again, this suggests
stored at Ϫ80°C. For analysis, a 50 l aliquot of the
that an N-oxide interferes with the NOS/NADPH stoi-
reaction mixture was directly injected into the HPLC
chiometry. It is known that NOϪ is able to oxidize
system without further sample processing. The HPLC
biological molecules (e.g., thiols and DNA), especially
system used was a Series 10 solvent delivery pump
in the presence of oxygen [21]. We examined therefore
(Perkin Elmer, Norwalk, CT, USA) equipped with an
the possible nonenzymatic redox reaction of NADPH
injection valve (Type 7125, Rheodyne, Cotati, CA,
with NOϪ and the effects of SOD, which converts NOϪ
USA) and connected to an UV detector operated at 230
to •NO. Furthermore, the amounts of the •NO decompo-
nm. A C-18 reverse phase chromatography column (5
sition products nitrite and nitrate were determined.
m particle size, 250 mm, 4.6 mm I.D.; CPS Analitica,Milano, Italy) was connected to a guard column (C-18, 5
EXPERIMENTAL PROCEDURES
m, 10 mm). The mobile phase contained 5 mM tetrabu-tylammonium hydrogensulfate and 10 mM Na HPO
and was adjusted to pH 6.4 with 0.2 M phosphoric acid.
NADPH was obtained from Applichem (Darmstadt,
The system was operated at a flow rate of 1.5 ml minϪ1.
Germany); Cu,Zn-SOD, from Roche Molecular Bio-
Calibration curves were prepared with standard solutions
chemicals (Penzberg, Germany). Angeli’s salt (AS) was
of potassium nitrite and nitrate. Chromatograms were
synthesized as published [21] and kindly provided by Dr.
recorded and processed with the use of an A C-R4A
David Wink (National Cancer Institute, Bethesda, MD,
integrator (Shimadzu Corporation, Kyoto, Japan).
USA). All other materials were of the highest purityavailable and obtained from either Sigma Chemicals
RESULTS AND DISCUSSION
(Deisenhofen, Germany) or Merck AG (Darmstadt, Ger-many). Water was deionized to 18 M⍀ cm (Milli-Q;
To investigate whether NOϪ, derived from AS, is able
Millipore, Eschborn, Germany) and de-oxygenated with
to oxidize NADPH, different concentrations of AS were
incubated together with 100 M NADPH and the reac-tion was followed at 340 nm (A
tions of AS and in the absence of SOD, 1 mol NADPHwas oxidized by about 4 moles AS (Fig. 1, open circles).
Angeli’s salt (AS) was dissolved freshly in 10 mM
At concentrations above 100 M AS, the relationship
NaOH; NADPH, in 0.5 M triethanolamine (TEA) buffer
between AS concentration and NADPH consumption
(pH 7.0). Aliquots of the Angeli’s salt stock solution
became nonlinear and reached a plateau (Fig. 1, inset),
were added to 94 well microtiter plates containing 100
most likely due to competing reactions of NOϪ, such as
M NADPH in 0.5 M TEA buffer (100 l). To controls,
dimerization yielding N O; however, the latter concen-
equal amounts of NaOH were added. The mixtures were
trations are about 10 times higher than product concen-
then incubated under protection from light in the absence
trations typically achieved in NOS activity assays.
or presence of 5 kU mlϪ1 SOD for 5 min at 37°C.
The stoichiometry of this reaction with respect to
Thereafter, the extinction was measured at 340 nm (A
NOϪ cannot be determined exactly, as the yield of NOϪ
with a SpectraMax 340 microplate reader (Molecular
from AS varies— depending on the presence of oxy-
gen— between 83 and 48% [24]. When the slight decom-
measure of NADPH oxidation, as this assay is commonly
position (12%) of the AS preparation used in this study
used to determine the redox state of NADPH, although
(see below) is taken into account, between 42 and 73
concurring reactions resulting in a similar loss in absor-
mol% NOϪ could have been formed per mol AS under
bance cannot be ruled out entirely. The results shown are
the investigated conditions. Thus, an initial reaction stoi-
representative of at least three individual experiments,
chiometry of 2 mol NOϪ per mol NADPH appears to be
When the nonenzymatic redox reaction of NADPH
Determination of nitrite and nitrate
with NOϪ is taken into account, the recently published
The NO-related metabolites, nitrite and nitrate, were
1:1.5 NADPH/NOϪ stoichiometry for H Bip-free NOS
determined as previously described [22,23] with the fol-
with N-hydroxy-L-arginine as substrate [20] may be
Fig. 1. NADPH oxidation by AS-derived NOϪ. NADPH (100 M) was incubated for 5 min at 37°C in the presence of differentconcentrations of the NOϪ-donor Angeli’s salt. Thereafter, the amount of oxidized NADPH was determined as described inExperimental Procedures. Open circles, reaction without SOD; closed circles, with addition of 5 kU mlϪ1 SOD. The dashed lineindicates an initial 1:2 stoichiometry of the NADPH oxidation with respect to NOϪ based on the considerations given in the text. Inset,NADPH oxidation at higher NOϪ concentrations. Data represent means Ϯ SEM (n ϭ 3–10 individual experiments, each performedin triplicate). An asterisk indicates statistical significance between the conditions with and without SOD (p Ͻ .01).
explained as follows: 1 mol NADPH would be consumed
H O formation or other uncoupled NOS reactions, al-
enzymatically; in addition, 0.5 mol would react directly
though they regarded this as unlikely under the condi-
with NOϪ to a yet-to-be-identified product (Fig. 2). The
tions applied. Our results now provide an alternative
authors of the aforementioned study suggested additional
explanation that does not imply such uncoupled NOS reac-tions. Recently, Stuehr’s laboratory found that NOS pro-duces different N-oxides depending on the presence ofH Bip [25], resulting of course in varying NADPH sto-
ichiometries. Thus the NADPH stoichiometry of the NOSreaction still remains a matter of debate and varies depend-ing on the availability of cofactors. The present studymight therefore be helpful in the interpretation of differ-ent experimentally observed NADPH/citrulline ratios.
Furthermore, we examined the effects of SOD, which
can convert NOϪ to •NO [12,26], on the oxidation ofNADPH by NOϪ. In this reaction system, SOD does notserve as a superoxide dismutating enzyme (under ourexperimental conditions, a source for superoxide canhardly be envisaged) but as a reactant in the reactionCu(II)SOD ϩ NOϪ N Cu(I)SOD ϩ •NO [13,26]. Asexpected, the addition of SOD (5 kU mlϪ1) markedlyprevented the oxidation of NADPH (Fig. 1, closed cir-cles). This is consistent with the rapid oxidation of NOϪ
Fig. 2. NOS-derived NOϪ. Different N-oxides generated by H Bip-free
by SOD to •NO, which by itself does not consume
NOS [20] from N-hydroxy-L-arginine and the corresponding NADPHstoichiometry.
Table 1. The Yield of Nitrite and Nitrate from the Reaction of
reactive nitrogen intermediate. That this reactive nitro-
Angeli’s Salt with NADPH and the Influence of SOD
gen species is ONOOϪ, generated by the reaction of
NOϪ with oxygen, is unlikely. ONOOϪ decomposes to
both nitrite and nitrate at a pH of 7.5 with a ratio of
approximately 1:2 [27]. The oxidation of NADPH by
ONOOϪ should result in the formation of nitrite, in
analogy to the oxidation of NADH by ONOOϪ [28]. Both the decomposition as well as the oxidation reaction
Angeli’s salt (100 M) was incubated with or without 100 M
NADPH in the absence or presence of 5 kU SOD mlϪ1 as described in
would thus have resulted in an increased level of either
Experimental Procedures. Thereafter, both NO metabolites were mea-
nitrite or nitrate, none of which was observed. The oxi-
sured by HPLC analysis. The inclusion of NADPH to the reaction
dation of NADPH by NOϪ therefore appears to be di-
mixture significantly decreased the amount of nitrite, whereas thepresence of SOD markedly increased nitrite levels. Nitrate levels were
rectly NOϪ-mediated. Candidate products for the metab-
almost one magnitude lower. Similar results have been obtained with
olites of NOϪ include the reduced form of the NOϪ
AS at concentrations of 200 and 400 M. The results represent the
dimerization product, hyponitrous acid, as well as hy-
means Ϯ SEM of n independent experiments. Student’s t-test wasperformed and p Ͻ .05 was considered to be statistically significant; *p
droxylamine (NH OH), a known product of both •NO
Ͻ .05 compared to control (AS, 100 M).
and NOϪ reduction [29 –31], or— by reaction with oxy-gen—a recently suggested derivative of ONOOϪ [32]. In
To examine possible products of the NADPH/NOϪ
the present study, we did not carry out anaerobic exper-
reaction, we determined the amounts of nitrite and nitrate
iments, as we focus on the relevance of our findings with
formed (Table 1). In the absence of NADPH, 100 M
respect to the aerobic NO synthase reaction. Neverthe-
AS (at a pH of 7.0) generated 88.3 Ϯ 9.4 M nitrite,
less, reinvestigating the reaction described here in the
while the yield of nitrate was as low as 5.3 Ϯ 0.6 M.
absence of oxygen would be of interest.
One mol of AS theoretically generates one mol of nitrite,
As nitrite levels were decreased when NADPH was
besides NOϪ. The yield of nitrite was, however, only
oxidized by NOϪ, the NADPH/NOϪ reaction product
0.88 mol per mol of AS, suggesting that approximately
might further react with nitrite. A possible reaction of
12% of the AS was decomposed. Similar values were
hydroxylamine with nitrite was therefore considered. Ni-
obtained at AS concentrations of 200 and 400 M (data
trite (100 M) was incubated with an equimolar amount
of hydroxylamine (100 M) for 5 min; thereafter, nitrite
In the presence of NADPH, the amount of nitrite
levels were determined by HPLC analysis as described
generated during AS decomposition was significantly
above. The presence of hydroxylamine however had no
lowered to an average of 66 Ϯ 5 M, whereas that of
influence on nitrite levels (99 Ϯ 6% of control); there-
nitrate remained almost unchanged (see Table 1). The
fore, the formation of hydroxylamine from NOϪ and
possibility exists that NOϪ reacts with NADPH via a
NADPH seems unlikely, although it cannot be ruled out
Fig. 3. AS-derived NOϪ. AS generates equimolar amounts of nitrite and NOϪ. The latter can dimerize yielding hyponitrous acid, whichdecomposes to nitrous oxide and water, or it can be converted to •NO by SOD. •NO can react with superoxide yielding peroxynitrite,which decomposes to nitrate and nitrite in a ratio of approximately 1:2. In the absence of superoxide, •NO decomposes predominantlyto nitrite. If NADPH is present, NOϪ will oxidize this cofactor. The metabolite of NOϪhas yet to be established; it appears to reactfurther with nitrite to a yet-to-be-identified reactive nitrogen species (confer also to [32]).
as the observed decrease of nitrite levels might well be
ysis in neuronal NO synthase. J. Biol. Chem. 274:24921–24929;
due to other side reactions. The definitive identification
[3] Griffith, O. W.; Stuehr, D. J. Nitric oxide synthases: properties
of the reduced form of NOϪ generated from the oxida-
and catalytic mechanism. Annu. Rev. Physiol. 57:707–736; 1995.
tion of NADPH thus warrants further investigation.
[4] Kwon, N. S.; Nathan, C. F.; Stuehr, D. J. Reduced biopterin as a
When SOD was present, the average nitrite content of
cofactor in the generation of nitrogen oxides by murine macro- phages. J. Biol. Chem. 264:20496 –20501; 1989.
the samples was almost doubled (Table 1). This is in
[5] Raman, C. S.; Li, H.; Martasek, P.; Kral, V.; Masters, B. S. S.;
accordance with the proposed conversion of NOϪ to NO
Poulos, T. L. Crystal structure of constitutive endothelial nitric
by SOD, as under the examined conditions, NO almost
oxide synthase: a paradigm for pterin function involving a novel metal center. Cell 95:939 –950; 1998.
completely decomposes to nitrite [33]. Figure 3 illus-
[6] Crane, B. R.; Arvai, A. S.; Gachhui, R.; Wu, C.; Ghosh, D. K.;
trates the different reaction pathways of NOϪ depending
Getzoff, E. D.; Stuehr, D. J.; Tainer, J. A. The structure of nitric
on the absence or presence of SOD and NADPH.
oxide synthase oxygenase domain and inhibitor complexes. Sci- ence 278:425– 431; 1997.
Under all conditions examined, nitrate levels ap-
[7] Crane, B.; Arvai, A.; Ghosh, D.; Wu, C.; Getzoff, E.; Stuehr,
peared to be one magnitude lower than nitrite levels. The
D. J.; Tainer, J. Structure of nitric oxide synthase oxygenase
increase in nitrate levels in the presence of SOD is most
dimer with pterin and substrate. Science 279:2121–2126; 1998.
[8] Marletta, M. A. Nitric oxide synthase structure and mechanism.
likely due to nitrate contaminations of the SOD prepa-
J. Biol. Chem. 268:12231–12234; 1993.
ration used: 5 kU mlϪ1 SOD alone contained 13 M
[9] Masters, B. S. S.; McMillan, K.; Sheta, E. A.; Nishimura, J. S.;
nitrate. Nitrate thus seems not to play a major role in the
Roman, L. J.; Martasek, P. Neuronal nitric oxide synthase, amodular enzyme formed by convergent evolution: structure stud-
tested reaction system. This is consistent with data from
ies of a cysteine thiolate-ligand heme protein that hydroxylates
neuronal cell cultures where the stimulation of NOS with
L-arginine to produce NO as a cellular signal. FASEB J. 10:552–
specific glutamate agonists or depolarizing agents gen-
erated always much higher amounts of nitrite than nitrate
[10] Mayer, B.; Klatt, P.; Werner, E. R.; Schmidt, K. Kinetics and
mechanism of tetrahydrobiopterin-induced oxidation of nitric ox-
[22]. The same situation was observed in a steady state
ide. J. Biol. Chem. 270:655– 659; 1995.
system like human cerebrospinal fluid in which the mea-
[11] Hobbs, A. J.; Fukuto, J. M.; Ignarro, L. J. Formation of free nitric
sured nitrite levels were four to five times higher than
oxide from L-arginine by nitric oxide synthase: direct enhance-ment of generation by superoxide dismutase. Proc. Natl. Acad.Sci. USA 91:10992–10996; 1994.
In conclusion, possible nonenzymatic side-reactions
[12] Schmidt, H. H. H. W.; Hofmann, H.; Schindler, U.; Shutenko,
of NOS-derived N-oxides with NADPH have to be taken
Z. S.; Cunningham, D. D.; Feelisch, M. No •NO from NO syn- thase. Proc. Natl. Acad. Sci. USA 93:14492–14497; 1996.
into account when the NADPH/L-citrulline stoichiome-
[13] Bahr, N.; Gu¨ller, R.; Reymond, J. L.; Lerner, A. L. A nitroxyl
try of NOS is determined. Although it might not be the
synthase catalytic antibody. J. Am. Chem. Soc. 118:3550 –3555;
major metabolic pathway of nitroxyl, our data highlight
[14] Xia, Y.; Zweier, J. Superoxide and peroxynitrite generation from
the oxidative potential of NOϪ in biological systems, as
inducible nitric oxide synthase in macrophages. Proc. Natl. Acad.
known for the oxidation of thiols— generating hydrox-
Sci. USA 94:6954 – 6958; 1997.
ylamine—and biological macromolecules [34], which
[15] Xia, Y.; Roman, L. J.; Masters, B. S. S.; Zweier, J. L. Inducible
nitric-oxide synthase generates superoxide from the reductase
might both compete with NADPH in the oxidative break-
domain. J. Biol. Chem. 273:22635–22639; 1998.
down of nitroxyl. Furthermore, we suggest a role of SOD
[16] Koppenol, W. H.; Moreno, J. J.; Pryor, W. A.; Ischiropoulos, H.;
in scavenging NOϪ, thus decreasing oxidative stress in
Beckman, J. S. Peroxynitrite, a cloaked oxidant formed by nitric oxide and superoxide. Chem. Res. Toxicol. 5:834 – 842; 1992.
cells. The importance of the apparently complex under-
[17] Stuehr, D. J.; Kwon, N. S.; Nathan, C. F.; Griffith, O. W.;
lying mechanism, with respect to the NOS reaction
Feldman, P. L.; Wiseman, J. N-hydroxy-L-arginine is an inter-
mechanism clearly warrants further investigations.
mediate in the biosynthesis of nitric oxide from L-arginine. J. Biol. Chem. 266:6259 – 6263; 1991.
[18] Mayer, B.; John, M.; Heinzel, B.; Werner, E. R.; Wachter, H.;
Acknowledgements — We thank Chiara Fenoglio for excellent exper-
Schultz, G.; Bo¨hme, E. Brain nitric oxide synthase is a biopterin-
imental assistance. We are indebted to Dr. David Wink for the generous
and flavin-containing multi-functional oxido-reductase. FEBS
gift of Angeli’s salt. L.Z. acknowledges the support from CARIPLO
Lett. 288:187–191; 1991.
Foundation (Milano) and from Associazione Italiana Parkinsoniani
[19] Chen, Y.; Rosazza, J. P. N. A bacterial nitric oxide synthase from
(Milano). This study was supported by the DFG (SFB 547/C7).
a Nocardia species. Biochem. Biophys. Res. Commun. 203:1251– 1258; 1994.
[20] Rusche, K. M.; Spiering, M. M.; Marletta, M. A. Reactions
catalyzed by tetrahydrobiopterin-free nitric oxide synthase. Bio-REFERENCES chemistry 37:15503–15512; 1998.
[1] Bo¨mmel, H. M.; Reif, A.; Fro¨hlich, L. G.; Frey, A.; Hofmann, H.;
[21] Wink, D.; Feelisch, M.; Fukuto, J.; Chistodoulou, D.; Jourd’heuil,
Marecak, D. M.; Groehn, V.; Kotsonis, P.; La, M.; Ko¨ster, S.;
D.; Grisham, M.; Vodovotz, Y.; Cook, J.; Krishna, M.; DeGraff,
Meinecke, M.; Bernhardt, M.; Weeger, M.; Ghisla, S.; Prestwich,
W.; Kim, S.; Gamson, J.; Mitchell, J. The cytotoxic mechanism of
G. D.; Pfleiderer, W.; Schmidt, H. H. H. W. Anti-pterins as tools
nitroxyl: possible implications for the pathophysiological role of
to characterize the function of tetrahydrobiopterin in NO syn-
NO. Arch. Biochem. Biophys. 351:66 –74; 1998.
thase. J. Biol. Chem. 273:33142–33149; 1998.
[22] Zecca, L.; Fariello, R. G.; Galimberti, M.; Racagni, G.; Ambro-
[2] Reif, A.; Fro¨hlich, L. G.; Kotsonis, P.; Frey, A.; Bo¨mmel, H. M.;
sini, A. Changes in nitric oxide metabolite levels in stimulated
Wink, D. A.; Pfleiderer, W.; Schmidt, H. H. H. W. Tetrahydro-
substantia nigra neurons. Neuroreport 8:2121–2125; 1997.
biopterin inhibits monomerization and is consumed during catal-
[23] Zecca, L.; Rosati, M.; Renella, R.; Galimberti, M.; Ambrosini, A.;
Fariello, R. G. Nitrite and nitrate levels in cerebrospinal fluid of
S-nitrosothiols and thiols: generation of nitroxyl (HNO) and sub-
normal subjects. J. Neural. Transm. 105:627– 633; 1998.
sequent chemistry. Biochemistry 37:5362–5371; 1998.
[24] Fukuto, J. M.; Hobbs, A. J.; Ignarro, L. J. Conversion of nitroxyl
[32] Ludwick, N.; Miranda, K. M.; Wink, D.; Fukuto, J. The physio-
(HNO) to nitric oxide (NO) in biological systems: the role of
logical chemistry of HNO. NO Biol. and Chem. 4:275; 2000.
physiological oxidants and relevance to the biological activity of
[33] Ignarro, L. J.; Fukuto, J. M.; Griscavage, J. M.; Rogers, N. E.;
HNO. Biochem. Biophys. Res. Commun. 196:707–713; 1993.
Byrns, R. E. Oxidation of nitric oxide in aqueous solution to
[25] Adak, S.; Wang, Q.; Stuehr, D. J. Arginine conversion to nitrox-
nitrite but not nitrate: comparison with enzymatically formed
ide by tetrahydrobiopterin-free neuronal nitric-oxide synthase:
nitric oxide from L-arginine. Proc. Natl. Acad. Sci. USA 90:8103–
implications for mechanism. J. Biol. Chem. 275:33554 –33561;
[34] Wink, D. A.; Kasprzak, K. S.; Maragos, C. M.; Elespuru, R. K.;
[26] Murphy, M. E.; Sies, H. Reversible conversion of nitroxyl anion
Misra, M.; Dunams, T. M.; Cebula, T. A.; Koch, W. H.; Andrews,
to nitric oxide by superoxide dismutase. Proc. Natl. Acad. Sci.
A. W.; Allen, J. S.; Keefer, L. K. DNA deamination ability and
USA 88:10860 –10864; 1991.
genotoxicity of nitric oxide and its progenitors. Science 254:
[27] Pfeiffer, S.; Gorren, A. C. F.; Schmidt, K.; Werner, E. R.; Han-
sert, B.; Bohle, D. S.; Mayer, B. Metabolic fate of peroxynitrite inaqueous solution. Reaction with nitric oxide and pH-dependentdecomposition to nitrite and oxygen in a 2:1 stoichiometry. ABBREVIATIONS J. Biol. Chem. 272:3465–3470; 1997.
[28] Kobayashi, K.; Miki, M.; Tagawa, S. Reactivity of peroxynitrite
as studied by pulse radiolysis. Endothelium 3(Suppl.):S65; 1995.
[29] Kroll, C.; Langner, A.; Borchert, H. H. Nitroxide metabolism in
H Bip—(6R)-5,6,7,8-tetrahydro-L– biopterin
the human keratinocyte cell line HaCaT. Free Radic. Biol. Med.
HPLC— high performance liquid chromatography
26:850 – 857; 1999.
[30] Kuppusamy, P.; Wang, P.; Zweier, J. L.; Krishna, M. C.; Mitchell,
J. B.; Ma, L.; Trimble, C. E.; Hsia, C. J. Electron paramagnetic
resonance imaging of rat heart with nitroxide and polynitroxyl-
albumin. Biochemistry 35:7051–7057; 1996.
[31] Wong, P. S.; Hyun, J.; Fukuto, J. M.; Shirota, F. N.; DeMaster,
ONOOϪ— oxoperoxonitrate(1Ϫ) (peroxynitrite)
E. G.; Shoeman, D. W.; Nagasawa, H. T. Reaction between
Oral Antibiotics Doxycycline, Minocycline, Tetracycline Oral antibiotics for acne have many more potential side effects. Our goal is to limit the dose and duration of oral antibiotic therapy as much as possible. Oral antibiotics used for acne can cause sunburn, antibiotic resistance, brain swelling and may also discolor baby’s teeth if used during pregnancy.
ERECTILE DYSFUNCTION Erectile dysfunction is an on- going inability to achieve or to maintain an erection sufficient for sexual intercourse. The likelihood of having erectile dysfunction increases with age. Anything interfering with blood flow to the penis can lead to erectile dysfunction; disease, injury or medication are common causes, as well as physical and psychological factors. Occa

 Free Radical Biology & Medicine, Vol. 30, No. 7, pp. 803– 808, 2001
Copyright 2001 Elsevier Science Inc.
Free Radical Biology & Medicine, Vol. 30, No. 7, pp. 803– 808, 2001
Copyright 2001 Elsevier Science Inc.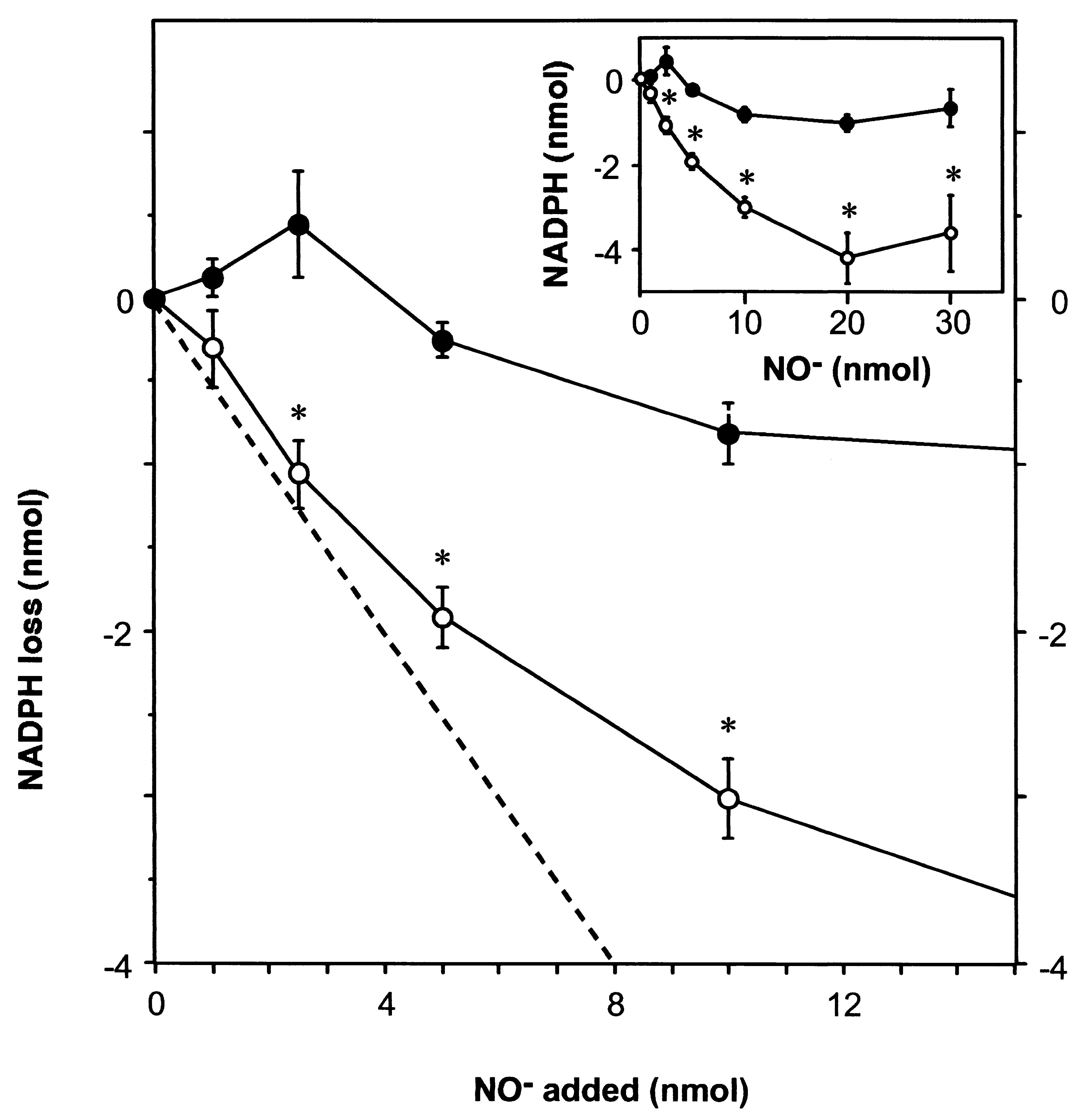
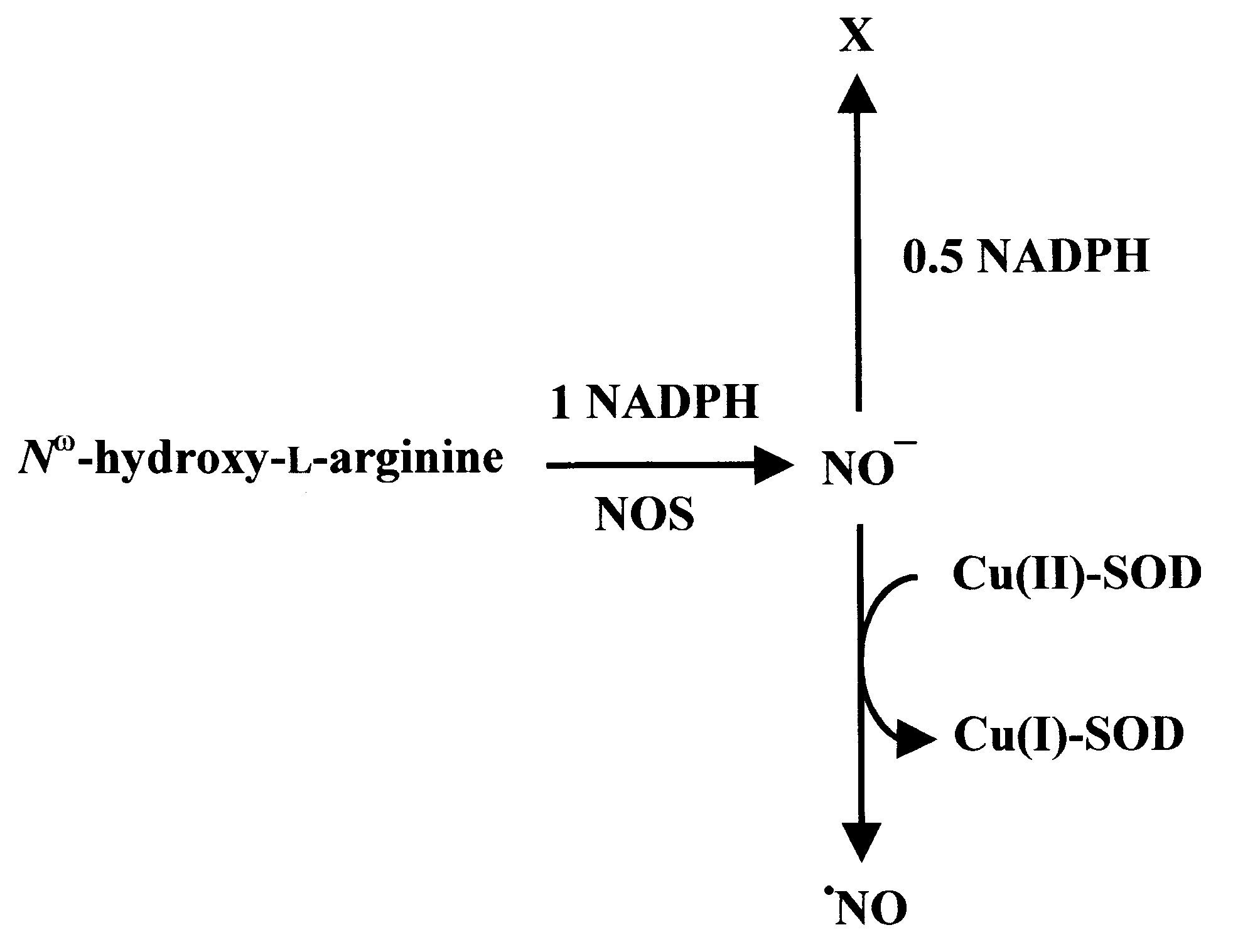 Fig. 1. NADPH oxidation by AS-derived NOϪ. NADPH (100 M) was incubated for 5 min at 37°C in the presence of differentconcentrations of the NOϪ-donor Angeli’s salt. Thereafter, the amount of oxidized NADPH was determined as described inExperimental Procedures. Open circles, reaction without SOD; closed circles, with addition of 5 kU mlϪ1 SOD. The dashed lineindicates an initial 1:2 stoichiometry of the NADPH oxidation with respect to NOϪ based on the considerations given in the text. Inset,NADPH oxidation at higher NOϪ concentrations. Data represent means Ϯ SEM (n ϭ 3–10 individual experiments, each performedin triplicate). An asterisk indicates statistical significance between the conditions with and without SOD (p Ͻ .01).
Fig. 1. NADPH oxidation by AS-derived NOϪ. NADPH (100 M) was incubated for 5 min at 37°C in the presence of differentconcentrations of the NOϪ-donor Angeli’s salt. Thereafter, the amount of oxidized NADPH was determined as described inExperimental Procedures. Open circles, reaction without SOD; closed circles, with addition of 5 kU mlϪ1 SOD. The dashed lineindicates an initial 1:2 stoichiometry of the NADPH oxidation with respect to NOϪ based on the considerations given in the text. Inset,NADPH oxidation at higher NOϪ concentrations. Data represent means Ϯ SEM (n ϭ 3–10 individual experiments, each performedin triplicate). An asterisk indicates statistical significance between the conditions with and without SOD (p Ͻ .01).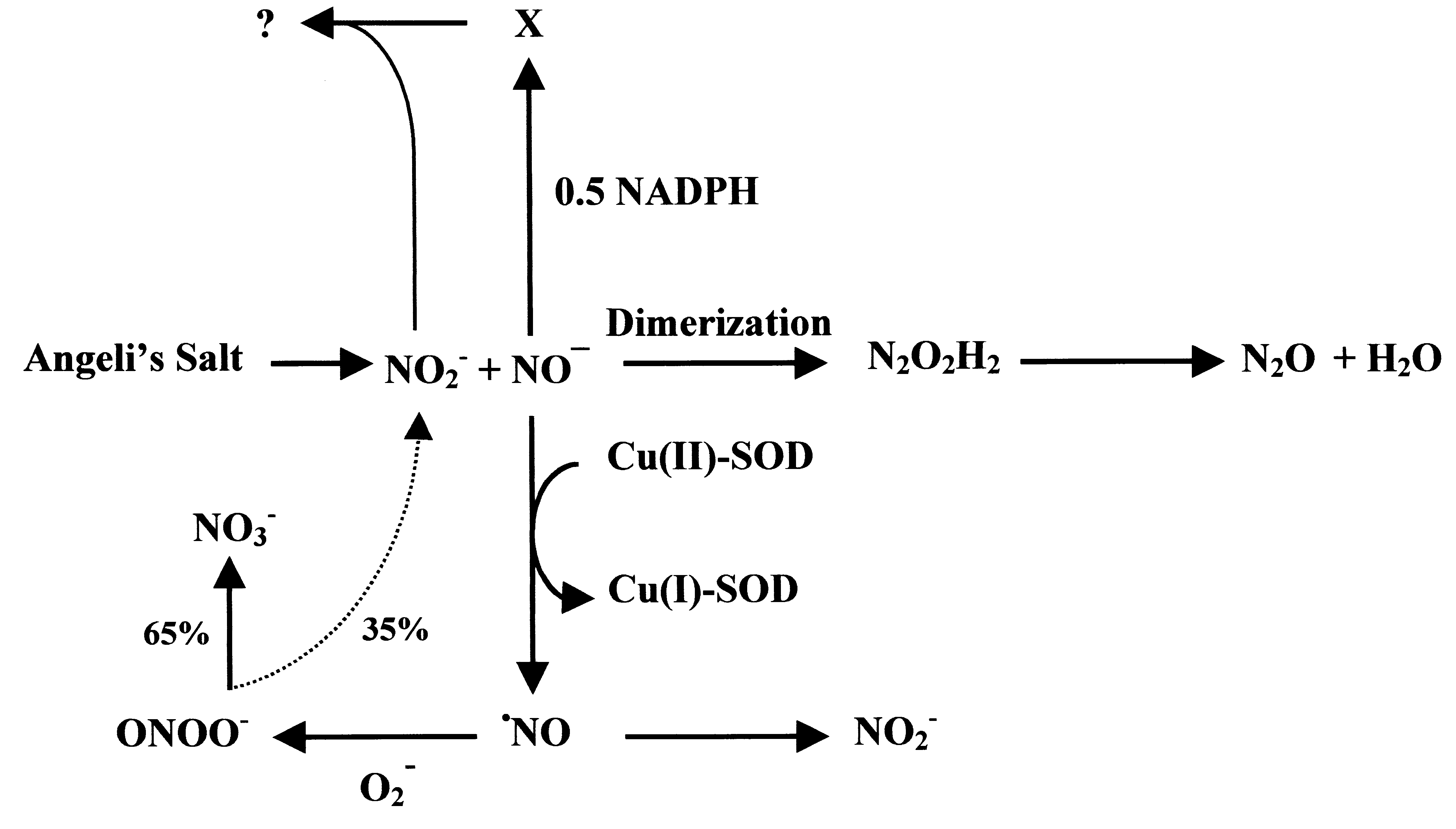 Table 1. The Yield of Nitrite and Nitrate from the Reaction of
reactive nitrogen intermediate. That this reactive nitro-
Angeli’s Salt with NADPH and the Influence of SOD
gen species is ONOOϪ, generated by the reaction of
NOϪ with oxygen, is unlikely. ONOOϪ decomposes to
both nitrite and nitrate at a pH of 7.5 with a ratio of
approximately 1:2 [27]. The oxidation of NADPH by
ONOOϪ should result in the formation of nitrite, in
analogy to the oxidation of NADH by ONOOϪ [28].
Table 1. The Yield of Nitrite and Nitrate from the Reaction of
reactive nitrogen intermediate. That this reactive nitro-
Angeli’s Salt with NADPH and the Influence of SOD
gen species is ONOOϪ, generated by the reaction of
NOϪ with oxygen, is unlikely. ONOOϪ decomposes to
both nitrite and nitrate at a pH of 7.5 with a ratio of
approximately 1:2 [27]. The oxidation of NADPH by
ONOOϪ should result in the formation of nitrite, in
analogy to the oxidation of NADH by ONOOϪ [28].