Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
hypothesis about the nature of electrodermal reactions
HYPOTHESIS ABOUT THE NATURE OF ELECTRODERMAL REACTIONS Dementienko V.V., Dorokhov V.B., Koreneva L.G., Markov A.G., Tarasov A.V. Shakhnarovitch V.M. Journal of Psychophysiology 1998.V.30/1-2, p.267.
Prolonged investigations on electrodermal activity (EDA) during activation - relaxation cycles,
including ones with simultaneous registration of resistance and temperature of the skin, were carried on. New facts observed (fast conductance decrease after its increase during electrodermal reaction - EDR, frequent absence of correlation between skin conductance and EDR frequency, the conditions of correlation between skin resistance and temperature) are difficult to explain by most known hypothesis concerning the formation of all EDA components by secretory part of eccrine sweat glands. They suggest the existence of two considerably independent systems of EDA regulation. According to the hypothesis, thermoregulation function is fulfilled mostly by continuous outflow of the secret , producing tonic EDA changes. EDR appear due to emotions and are poorly connected to thermoregulation. They are produced by excretory parts of the glands; the resistance drop corresponds to the duct contraction caused by mioepitelium sells under the influence of activation of sympathetic system, probably adrenergic.
INTRODUCTION Electrodermal activity (EDA), or the change of skin electric properties, is one of the most known,
(Aldersons 1985, Dawson 1990, Edelberg 1971, Prokasy 1973) and widely used, (Collet 1996, De Pascalis 1995, Vossel 1984, Lim 1996) electrophysiological parameters. It is well-established (Edelberg 1971, Dawson 1990, Freedman 1994) to be connected to the sweat excretion by eccrine glands (MacGoodall 1970, Robertshow 1977, Sato 1983, Wolf 1974, Uno 1977), whose activity is regulated by sudomotor neurons of sympathetic system (MacGoodall 1970 , Sato 1983, Uno 1977 Lidberg 1981, Shields 1987). Main function of sweating is universally recognized to be the body cooling during heat or work. It is also known that after stimuli electrodermal responses (EDR) appear, in which significant skin conductance changes during seconds are observed. EDR are also called phasic EDA component, as differing from tonic one, when changes comparable to dynamic range appear in minutes (Lovibond 1992). EDR without registered stimuli are called nonspecific or spontaneous.
In spite of universally recognition of partition of EDA into tonic and phasic components, and its
genesis - into thermoregulational and emotional, both partitions are not done successively. Function of emotional EDA is considered to be the additional cooling, and often any resistance change of appropriate value following the stimuli is considered "response", see for example Vossel 1984. All peculiarities of EDA are attributed to the way of activity of neurons innervating secretory part of the gland. The functions of cholinergic and adrenergic neurons are considered similar.
We don't believe in identical nature of activity components differing in times of considerable changes
for about two orders. Phasic components could have a separate function and somewhat independent regulation connected at least partly with adrenergic innervation of eccrine glands (Uno 1977, Shields 1987). Hypothesis about "double regulation" of sweating had been proposed in connection with "emotional sweating" (Bligh 1967), but later was rejected, see below.
The aim of the work is to show that the most popular hypothesis on the nature and functions of EDR
could not explain facts and that reconsidered and modified hypothesis of “double regulation” could explain them better.
For substantiation of the hypothesis about the nature and functions of EDR, we have carried on
additional experiments. Main differences of our experiments from previous ones known to us are as follows. 1) Experiments included several activation-relaxation-drowsiness cycles, 2) EDR are recognized more accurately, 3) along with sudomotor activity, a vasomotor one, also participating in thermoregulation and reacting on the same stimuli, was investigated.
METHODS Three independent measures of skin resistance with external voltage source 0.9 v. were taken. As
electrodes, two pairs of rings on fingers and the bracelet on the left wrist were used. Area of the electrodes 1
cm . The registration differed from that recommended (Fowles 1981) in using “dry” electrodes. Control experiments show that the difference through using such electrodes is actual only during first 5 minutes, which was not important to our experiments, see below.
In addition the following electrophysiological parameters were recorded: 2 monopolar EOG (horizontal
and vertical) using left mastoid reference, monopolar EEG (С ) using right mastoid reference, ECG
(nonstandard – left arm - left mastoid) and the voltage of the diod due to pressing. For registration, 8-channel polygraph Maclab 8.1 connected to Macintosh computer was used. The sampling rate 100 Hz, with 12 bit ADC. The amplifier bandwidths for EOG, ECG and one of EDA channels were from 0.7 (time constant 1.5 s.) to 20 Hz, for EEG - 0.7-50 Hz., for two EDA channels – high frequency filters (upper limit 20 Hz).
The channel with band-pass filter allows to observe EDR, but distorts them, therefore the main method
of EDR registration was recognition from the data obtained without the filter. For this, special software based on the form of the dependence of conduction on time was used, see below.
Temperature of the room was 21-24 degrees. Vasomotor activity was studied by means of dynamical IR -thermography (Anbar 1994, Gulyaev 1987,
1995) Thermograms of hands were recorded simultaneously with skin resistance monitoring by means of separate computer .
108 voluntiers aged from 16 to 69 with good alpha-activity in EEG without sleep deprivation were the
subjects of experiments. They sat in comfortable posture. The hands were put on the support without fixation. During thermographic experiments the subjects were asked not to move at least one hand. Skin resistance was continuously monitored for no less than 40 minutes, skin temperature – about 30 min. Most subjects participated in several sessions. At the beginning the subjects were activated by means of mental or emotional stress (mental calculations, unpleasant talk, computer games when unsuccessful, unpleasant sounds). Two kinds of experiments were undertaken. In experiments of the first type, the subjects were oriented on maintaining awake and active. They sat with open eyes in illuminated room and could talk with one another or with experimenter, read books, play computer games successively. In such experiments participated 43 subjects, 20 men and 23 women. In experiments of another kind, the orientation was given to decrease of alertness, up to going asleep. The subjects sat with closed eyes in a dark room and performed a monotonous psychomotoric test: pressed the button of piezodiode while counting from 1 to 10, then counted to 5 without pressing the button and so on. Performance of the test did not change significantly during first stages of drowsiness; when passing to more profound drowsiness errors occurred, when falling asleep the pressings became accidental or stopped. After 15 minutes the room was illuminated, the subjects opened eyes, and the experimenter activated them. In this type of experiment, 64 subjects participated, 29 men and 35 women.
12 experiments of the first type and 11 – of the second were undertaken without previous activation. For statistical treatment of the results, software STATISTICA 6 was used.
RESULTS For investigations of the EDR shape we used single responses registered 1-5 s after known stimuli
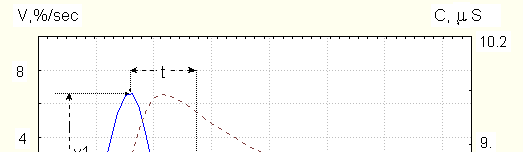
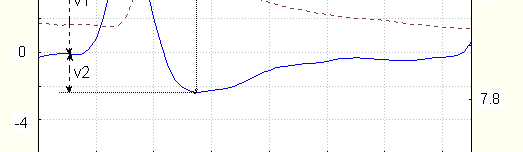
(breath inhalation, sharp sound). Its peculiarities are seen on Fig. 1, on which time dependence of conductance and its derivative by time are shown.
Dotted line is conductance in S (1 S= 1/1Om), right axis, solid line – time derivative in
%/sec. (left axis). V1,V2 are maximal velocities of conductance increase and decrease,
assimetry is v1/v2. (For this example v1/v2=2.7); t is the time of most changes (between
• the existence of fast conductance decrease after its initial increase;
• asymmetry – the ratio of the maximal speed of conductance increase to that of decrease is more than 1;
• time parameter of most significant changes is less than 6 sec.
The time of conductance increase is 1-3 s, its maximal speed during single EDR (derivative maximum)
- up to 15% per sec. see also Dawson 1990, the time of fast conductance decrease is no more than 3-5 s. Frequently, the signals with small asymmetry (1.2-1.3) are seen ; for several subjects during active awakeness such signals prevail. In case of almost symmetrical signals, the time of fast conductance decrease could be no more than 1.5-2 s. After fast conductance decrease, its more slow decrease is seen.
Intervals between EDR (between sequential derivative minimums) were determined by special
software. When the subjects are awake and active, 85% of intervals are between 5 and 60 s. When the alertness decreases, as seen by EEG, EOG, ECG and performance, the finger skin resistance increases, see also Lim, 1996. EDR frequency decreases; for 92 subjects during experiments of both types the alertness decrease is accompanied by EDR disappearance for several minutes. The subject's condition at the time of last EDR corresponds to slight drowsiness with small changes in EOG and without performance errors. The transition to real drowsiness (as`seen by EEG, EOG, ECG and errors) could be observed no less than a minute after last EDR.
If spontaneous EDR were not observed for more than two minutes, usual stimuli (breath inhaling,
sounds, unexpected touching) didn't provoke EDR. Activation beginning is followed by resistance decrease,
mostly with less speed than during EDR, without the appearance of EDR with appropriate form. After this , EDR begin to appear, at first in response to stimules (stimulated), then without registered stimulus (spontaneous). The example of skin resistance change during transition to drowsiness and activation is seen on Fig.2.
The example of resistance changes during cycles (solid). Vertical lines (dotted) show the position of EDR as found by software. Arrows above: a1, a2 –activation, r1,r2- relaxation.
maintained awakeness could be considered the succession of activation-relaxation periods, with peculiarities similar to those described earlier. In such conditions however in more than half experiments relaxation is not prolonged enough for entire EDR disappearance, and when conductance increases due to "activation", EDR appear immediately.
Two more rare cases must be mentioned: complete EDR absence during the experiment (recorded 22
times, in 18 of which there was no activation at the beginning) and continuous EDR outflow with all intervals in the region 5-10 s (15 times). In such cases resistance level remains constant or decreases.
The wrist resistance change could be described in the same way as that of the fingers. If EDR are
registered, all processes on hands and wrists are almost synchronous (crosscorelation coefficients are more than 0.8). However, for the wrist EDR absence is seen considerably more often than for fingers (in more than 50% of experiments - for all their duration). Tonic resistance decrease is frequently seen, and activation periods are notable by decrease acceleration.
The trend of conduction changes often doesn't depend on EDR existence and frequency. The cases of
conductivity level increase without EDR, more often observed on the wrist, were already mentioned. To the contrary, the decreasing or constant conductivity level is often observed along with numerous EDR, in particular following each other with minimal intervals 5-10 s.
Statistically significant correlation (of the order 0.2-0.3) between the current resistance level and
interval between EDR is seen only in about 50% of experiments of both types. In the rest of experiments correlation mentioned, as well as correlation between conductance level and average EDR intensity, is absent. Correlation between average conductance level and average EDR frequency is also absent. For
example, for three subjects resistance level approached maximal one for all experiments, and EDR frequency was above average, and for 5 subjects both resistance level and EDR frequency were below average.
Strong (up to 0.8) correlation is observed between average conductivity level and temperature.
Correlation between EDR frequency and temperature is significantly less.
When fingers skin resistance and hand temperature were measured simultaneously, significant
crosscorrelation between them was often found : the periods of temperature increase and decrease coincide with those of resistance. Crosscorrelation coefficients could be as high as 0.8.The example is seen in Fig. 3
Resistance is solid line, left scale, temperature –dotted one, right scale.
Correlation is maximal if during resistance changes several EDR are observed. It is absent if only tonic
resistance changes are seen. In particular, it disappears during relaxation with EDR disappearance, and reappears with their beginning after activation. That is why the said correlation is found less frequently in experiments of
Resistance – solid line, temperature –dotted.
Correlation is significantly less, especially when - drowsiness and sleep are present (500-1000 and
If EDA activation is obtained by transference of the subject to the room with higher temperature, (up to
27 degrees) , the resistance decreases (tonically), while the temperature increases, as should be expected for thermoregulation (Hensel 1981), that is resistance and temperature change in opposite directions. Then the temperature and conductance increase stops, and numerous EDR could be seen, see Fig.5.
Opposite trends of temperature and resistance are also observed for persons that systematically take the
medicines blocking the adrenoreceptors activity (dihydropyirydin derivatives- nifedipine and the like). It had been shown that when certain blockators concentration is achieved, the usual stimuli activating sympathetic nervous system evoke hands temperature increase instead of its usual decrease (Gulyaev 1994). In conditions mentioned, both stimulated and spontaneous EDR are absent, and stimuli cause tonic resistance decrease (seen no less than 20 times).
DISCUSSION As a result of our investigations the following EDA features were recognized. 1) EDR has a shape with
a substantial region of negative derivative, so that the velocities of conductance increase and decrease could have similar values (from 1 to 30 % in 2-3 s). Attempts had been taken to connect fast conductance decrease with psychological factors (Prokasy 1973). Later however the shape of EDR signals didn't draw proper attention. . 2) Correlation between skin resistance, its tonic change and overall intensity of electrodermal reactions is absent in more than half experiments, especially in ones when activation-drowsiness cycles were studied. Therefore, skin resistance and its changes appear to be not always determined by EDR. Low levels of correlation between phasic and tonic EDA are known,. (Freedman 1994, Lim 1996), but the fact seems to be often neglected. 3) Strong correlation between skin resistance and temperature dynamics is observed when changes are due to emotions, leading to EDR development. In conditions of thermal load, and under adrenergic block, resistance and temperature change in opposite directions.
We believe those facts couldn't be explained using most universally recognized hypothesis on EDR
Main features of the most recognized hypothesis: EDR as well as tonic EDA changes are connected
with processes in secretory part of the sweat gland and are regulated by the same neurons, primary cholinergic postgangleous neurons of sympathetic system; all peculiarities of EDR are connected to the way of activity of neurons innervating this part.
Main features of hypothesis under consideration: phasic and tonic EDA are formed by different parts of
the gland and have at least partially independent neurogenic regulation, possible different central drives (Mangina, 1996); phasic EDR are formed in excretory part of the gland; the system regulating EDR could be adrenergetic; EDR could have additional function.
Let us consider in more detail the formation of EDR according to both hypotheses. We have not seen in references the detailed description of EDR formation according to the first
hypothesis. EDR are connected to the secretion of the sweat portion as a response to stimulus activating sympathetic nervous system. The portion is thrown on skin surface through the excretory part opening passively. The following resistance increase is connected to sweat spreading or its readsorbtion by the skin.
The second hypothesis is as follows. Sweat, as in recognized one, is produced in secretory part.
Production and exit to the surface occur during minutes and determine skin resistance level and its tonic changes. EDR however are produced in excretory part of the gland in response to activating stimuli. Due to constriction of mioepitelium cells surrounding the part, the secret produced in secretory part and partly filling the duct, raises to skin surface. The resistance decreases due to appearance of additional contacts with electrodes. Following fast resistance increase is connected with braking of contacts due to elastic
mioepithelium dilatation after the stimuli disappearance.
Obviously, the duct constriction will cause EDR only on condition that enough secret is produced in
secretory part. Therefore any influence reducing secretory function decrease also pulse function. However, sometimes the EDR will not appear even when there is enough secret, if the regulation of excretory part is al least partly independent of that of the secretory one.
The hypothesis concerning the possibility of sweat formation in response to emotional stimuli by
mioepithelium of secretory part, was formulated previously (Bligh 1967). It was discussed also in connection with EDR (see reviews, for example Aldersons 1985), but was rejected. The main reasons of the rejection are as follows. 1) Emotional sweating is known to appear without mioepithelium constriction, 2) There is little amount of secret in excretory part, so its constriction can't produce much sweat (MacGoodall 1970, Sato 1983) , 3) Such possibility contradicts some of mioepithelium properties in vitro (Sato 1980). The hypothesis was not reconsidered later.
The reconsidered hypothesis explains not "emotional sweating" but EDR. In response to stimuli tonic
decrease of resistance could occur, followed by sweating, but not EDR. For EDR appearance much sweat excretion isn't necessary. The constriction followed by raise of the secret up to reaching the electrodes is enough. The conduction increases due to the increase of the number of ducts contacting the electrodes. ("active" ). Due to single EDR this number can increase by several percent - conductance increases by the same order. In favor of the hypothesis is the fact of the significant increase of EDR amplitude in case of artificial rise of the sweat level in the duct (Edelberg 1983).
We believe the hypothesis under consideration does not contradict the known mioepithelium properties.
Pulsatory activity of mioepithelium of human and animal apocrine glands under continuos catheholamine influences is shown. For eccrine glands, more precise regulation could take place. Mioepithelium of these glands in vitro is known to be inactive under adrenaline application, and continuous acetilholin application doesn't produce pulsatory activation (Sato 1980). That doesn't however contradict the supposition that it could be produced by pulsatory activity of neurogenous noradrenaline. Such activity of hand skin neurons in response to stimuli is registered; it correlates with EDR appearance (Lidberg 1981). Innervation of excretory parts of eccrine glands is also demonstrated (Kennedy,1994).
Let us consider the results described here according to both hypotheses. The shape of EDR is very difficult to explain according to the most recognized hypothesis. Firstly, the
secretion in several seconds hadn't been described. Secondly, it is hard to explain the regulating impulse that could drive full secretion of sweet portion through passive duct in 1-2 seconds. Practically impossible is to explain by passive sweet spreading or reabsorption the phase of swift conductance decrease, especially when times of conductance increase and decrease are of the same magnitude.
To the contrary, in supposition that times of conductance changes through EDR are connected with
elastic properties of mioepithelium, the features of EDR common in all experiments, could be readily explained. The known elastic properties of mioepithelium are similar to those of vessel smooth muscles. (Dudel, 1981) The constriction of contracting parts under the influence of nervous impulse is known to take 1-3 sec., the elastic relaxation - 3-5 sec. (Sato, 1980). The times of raise (relaxation) of the secret are similar. The following passive readsorbtion produces slow resistance increase
Frequently enough absence of correlation between EDR intensity and skin conductance and its trend
is easily explained in assumption of partly independent regulation of sweat secretion and EDR generation. Correlation absence corresponds to the cases when the secretion system is more excited than the EDR generation one. To explain these facts according to the well known hypothesis one must suppose either the existence of two gland populations (not confirmed morphologically), or of two ways of secretory part working (pulsate and continuos), observed simultaneously.
The resistance changes in wake-drowsy cycles is also explained easily by the second hypothesis.
According to it, the cycles of resistance correspond to continuous changes of secretory function. EDR disappear when the excitation of "emotional" function decreases, and do not reappear while there is not enough secret . Producing of it (and passive outflow to the surface) corresponds to tonic resistance decrease
during activation. To the contrary, if there is only one regulation region, one must suppose that the mode of regulation changes two times during the cycle, and the times of the said changes don't coincide with those of other electrophysiological parameters (EEG, EOG, ECG). Such supposition at least requires additional explanation.
In favor of hypothesis of independent regulation is also the fact of existence of correlation of resistance
and temperature dynamics in EDR presence and its absence for tonic EDA.
The existence of correlation of vaso- and sudomotor activity in EDR presence suggests that EDR
formation could be connected to adrenergic part of sympathetic system that is known to determine vasomotor activity. (Dudel 1980) The additional argument is the fact found by us that EDR, but not sweating, disappear under adrenergetic blockade. Finaly, in favor of it is the strong resemblance of EDR produced by eccrine glands and the work of apocrine glands, for which the adrenergic regulation of pulsate activity is proved.( MacGoodall 1970, Uno 1977).
The suggested function could be attributed to the neuron population that haven’t changed mediator
during development (Habecker 1994). It could be the neurons that while being sudomotor and not vasomotor influence cardiovascular system; such neurons had been reviled by direct measurements (Macefield 1996). The density of adrenergic synapses is the most on palms, but they are found also on the wrist, (Uno 1977), where EDR are also observed. The neurons under consideration could have axons innervating secretory parts of the eccrine glands (Kennedy 1994) producing the tonic resistance changes due to emotions , observed extremely often on wrists. The existence of axons going along the duct is shown (Kennedy 1994) .
The second, adrenergic regulation system could be supposed to have independent function. The
purpose of "emotional" EDR could be to inform other persons on the condition of a given person - for example on the degree of fear, threat or pleasure. The information could be contained in EDR intensity and frequency.
The increase of EDR frequency on temperature elevation could suggest that the discussed system have
thermoregulating function, as was supposed earlier. We believe however that the said transition to succession of EDR with small intervals could be due to the fact that significant temperature increase is very unpleasant to most people. In support of this suggestion is the fact that for the three persons to whom the elevation of room temperature to 28 degrees didn't seem unpleasant, the rapid EDR didn't appear; the resistance decreased as for tonic EDA. To the contrary, rapid EDR successions were easily produced for all subjects by extremely unpleasant for them influences, such as unexpected loud sounds, unpleasant words and so on.
SUMMARY New facts observed (fast conductance decrease after its increase during electrodermal reaction - EDR,
frequent absence of correlation between skin conductance and EDR frequency, the conditions of correlation between skin resistance and temperature) are difficult to explain by most known hypothesis concerning the formation of all EDA components by secretory part of eccrine sweat glands. They suggest the existence of two considerably independent systems of EDA regulation. According to the hypothesis, thermoregulation function is fulfilled mostly by continuous outflow of the secret , producing tonic EDA changes. EDR appear due to emotions and are poorly connected to thermoregulation. They are produced by excretory parts of the glands; the resistance drop corresponds to the duct contraction caused by mioepitelium sells under the influence of activation of simpatico system, probably adrenergic.
REFERENCES Aldersons A.A. The nature of electrodermal reactions. (Rus). 1985. Riga, Zinatie, 129 p Anbar M. Quantitative dynamic telethermometry in medical diagnosis and management. CRC Press,
Bligh J.A. A thesis concerning the processes of secretion and discharge of sweat. Evironm. Res. 1967,
Collet C., Roure-R., Rada H e.a. Relationships between performance and skin resistance evolution
involving various motor skills. // Physiol.Behav. 1996 V 59 N4 P. 953
Dawson M.E., Shell A.M., Filion D.N. The electrodermal system. In: Principles of psychophysiology
(J.T. Cacioppo, L.G. Tassinary Ed.) Cambridge Univ. Press, 1990, p. 295-324.,
Dudel.J, Rueg I., Shmidt R., Yanig V. Human physiology. Blood, bloodflow. 290 p. Edelberg. R Electrical properties of the skin. In: Biophysical properties of the skin. (H.R. Elden ed.)
Edelberg R. The effect of initial levels of sweat duct filling and skin hydration on electrodermal
response amplitude. // Psychophysiology 1983, V.20 N5 P. 550
Freedman L.W,.Serbo A.S, Dawson M.E. e.a. The relationship of sweat glands count to electrodermal
activity. //Psychophysiology 1994, V.31 N 2 P.196
Fowles. D.C,.Chritie M.J,.Edelberg R e.a. Publication recommendations for electrodermal
measurements. Psychophysiology 1981, V.18 N2 P. 232
MacGoodall C. Innervation and inhibition of eccrine and apocrine glands. J. Clinical Pharmacology,
Gulyaev Yu.V., Godik E.E., Markov A.G. et al. Infrared dynamical thermography of biological
objects. // Intl. J. Infrared Millim. Waves 1987 V. 8 N5 P. 317.
Gulyaev Yu.V., Markov A.G., Koreneva L.G, Zakharov P.V. Dynamical Infrared Thermography in
Humans.// IEEE. E M B. 1995 V. 14, N 6, , P.766
Habecker B.A., Landis S.C. Noradrenergic regulation of cholinergic differentiation. Science. 1994 V
Hensel H. Thermoreception and temperature regulation. N-Y, Academic. 1981 321 p. Kennedy W.R., Wendelschafer C.G., Brelje T.C. Innervation and vasculature of human sweat glands:
an immunohistochemistry-laser scanning confoal fluorescence microscopy study.// J.Neurosci. 1994 V 14 N11 Pt 2 P. 6825
Lidberg L,.Wally B.G. Sympathetic skin nerve discharges in relation to amplitude of skin resistance
responses.// Psychophysiology 1981, V.18 N2 P. 268
Lim C.L., Barry R.J., Gordon E., e.a. The relationship between quantified EEG and skin conductance
level. Int. J. Psychophysiol. 1996, 21(2-3) p. 151-62
Lovibond P.F. Tonic and phasic electrodermal measurements of human aversive conditioning with long
duration stimuli. Psychophysiology 1992, v.29 N 6 p. 621-633
Mangina C.A., Beuzeron-Mangina J.H. Direct electrical stimulation of specific human brain structures
and bilateral electrodermal activity.// Int-J-Psychophysiol. 1996 V. 22 N1 P. 1
Macefield V.G., Wallin B.G. The discharge behaviour of single sympathetic neurones supplying human
sweat glands.// J.Auton.Nerv.Syst. 1996 V.61N3 P. 277.
De Pascalis V., Barry R. J., Sparita A. Decelerative changes in heart rate during recognition of
visual stimuli: effects of psychological stress. Int. J. Psychophysiol. 1995 20(1)p. 21-31
W.F.Prokasy ed. Electrodermal activity in psychological research. NY, Acad.,1973. Robertshow D. Neuroendocrine control of sweat glands.// J. Invest. Dermat. 1977, V. 69 N1, P. 121. Sato. K. The physiology and pharmacology of the eccrine sweat gland. In: Biohemistry and physiology
of the skin, (L.A. Goldsmith ed.) N.Y. 1983, p. 596-641
. Sato K. Pharmacological responsiveness of the myoepithelium of the isolated human axillary apocrine
sweat gland. // Brit.J. Dermat., 1980, V.103 N3, P. 235.
Shields S.A,.MacDowell K.A,.Fairchild S,.Campbell M.L. Is mediation of sweating cholinergic,
adrenergic or both. // Psychophysiology 1987, V.24 N 3, P.312.
Uno H. Sympathetic innervation of the sweat glands and piloerrector muscles of macaques and human
beings.// J. Invest. Dermat. 1977, V. 69 N1, P. 112.
Vossel G., Kossman R. Electrodermal habituation speed and visual monitoring performance.
// Psychophysiology 1984, V. 21 N 1, P.97.
Wolf J.E,. Maibach H.I. Palmar eccrine sweating - the role of adrenergic and cholinergic mediation.
// Brit. J. Dermat., 1974, V.1 N4, P. 439.
Effects of calcium alkaline ionized water on formation and maintenance of osseous tissues (osteoporosis). Rei Takahashi, Zhenhua Zhang, Yoshinori ItokawaKyoto University Graduate School of Medicine, Dept. of Pathology and Tumor Biology, Fukui Prefectural University, Japan. "Effects of calcium alkaline ionized water on formation and maintenance of osseous tissues in rats were examined.
When (Not) to Stop a Clinical Trial for Benefit JAMA . 2005;294(17):2228-2230 (doi:10.1001/jama.294.17.2228) Medical Practice; Medical Ethics; Randomized Controlled Trial Randomized Trials Stopped Early for Benefit: A Systematic Review 21. Alliance for Cervical Cancer Prevention. Effectiveness, safety, and acceptabil- 25. Crum CP. The beginning of the end for cervical cancer? N Engl
 RESULTS
RESULTS 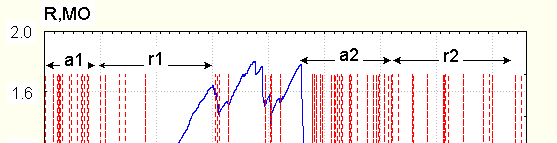
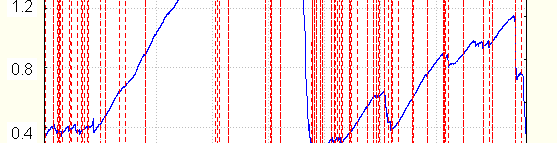
 mostly with less speed than during EDR, without the appearance of EDR with appropriate form. After this , EDR begin to appear, at first in response to stimules (stimulated), then without registered stimulus (spontaneous). The example of skin resistance change during transition to drowsiness and activation is seen on Fig.2.
The example of resistance changes during cycles (solid). Vertical lines (dotted) show the position of EDR as found by software. Arrows above: a1, a2 –activation, r1,r2- relaxation.
maintained awakeness could be considered the succession of activation-relaxation periods, with peculiarities similar to those described earlier. In such conditions however in more than half experiments relaxation is not prolonged enough for entire EDR disappearance, and when conductance increases due to "activation", EDR appear immediately.
Two more rare cases must be mentioned: complete EDR absence during the experiment (recorded 22
times, in 18 of which there was no activation at the beginning) and continuous EDR outflow with all intervals in the region 5-10 s (15 times). In such cases resistance level remains constant or decreases.
The wrist resistance change could be described in the same way as that of the fingers. If EDR are
registered, all processes on hands and wrists are almost synchronous (crosscorelation coefficients are more than 0.8). However, for the wrist EDR absence is seen considerably more often than for fingers (in more than 50% of experiments - for all their duration). Tonic resistance decrease is frequently seen, and activation periods are notable by decrease acceleration.
The trend of conduction changes often doesn't depend on EDR existence and frequency. The cases of
conductivity level increase without EDR, more often observed on the wrist, were already mentioned. To the contrary, the decreasing or constant conductivity level is often observed along with numerous EDR, in particular following each other with minimal intervals 5-10 s.
Statistically significant correlation (of the order 0.2-0.3) between the current resistance level and
interval between EDR is seen only in about 50% of experiments of both types. In the rest of experiments correlation mentioned, as well as correlation between conductance level and average EDR intensity, is absent. Correlation between average conductance level and average EDR frequency is also absent. For
mostly with less speed than during EDR, without the appearance of EDR with appropriate form. After this , EDR begin to appear, at first in response to stimules (stimulated), then without registered stimulus (spontaneous). The example of skin resistance change during transition to drowsiness and activation is seen on Fig.2.
The example of resistance changes during cycles (solid). Vertical lines (dotted) show the position of EDR as found by software. Arrows above: a1, a2 –activation, r1,r2- relaxation.
maintained awakeness could be considered the succession of activation-relaxation periods, with peculiarities similar to those described earlier. In such conditions however in more than half experiments relaxation is not prolonged enough for entire EDR disappearance, and when conductance increases due to "activation", EDR appear immediately.
Two more rare cases must be mentioned: complete EDR absence during the experiment (recorded 22
times, in 18 of which there was no activation at the beginning) and continuous EDR outflow with all intervals in the region 5-10 s (15 times). In such cases resistance level remains constant or decreases.
The wrist resistance change could be described in the same way as that of the fingers. If EDR are
registered, all processes on hands and wrists are almost synchronous (crosscorelation coefficients are more than 0.8). However, for the wrist EDR absence is seen considerably more often than for fingers (in more than 50% of experiments - for all their duration). Tonic resistance decrease is frequently seen, and activation periods are notable by decrease acceleration.
The trend of conduction changes often doesn't depend on EDR existence and frequency. The cases of
conductivity level increase without EDR, more often observed on the wrist, were already mentioned. To the contrary, the decreasing or constant conductivity level is often observed along with numerous EDR, in particular following each other with minimal intervals 5-10 s.
Statistically significant correlation (of the order 0.2-0.3) between the current resistance level and
interval between EDR is seen only in about 50% of experiments of both types. In the rest of experiments correlation mentioned, as well as correlation between conductance level and average EDR intensity, is absent. Correlation between average conductance level and average EDR frequency is also absent. For
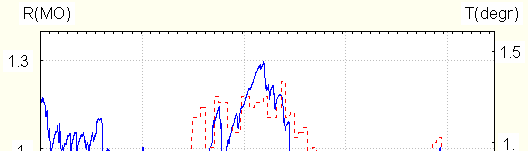
 example, for three subjects resistance level approached maximal one for all experiments, and EDR frequency was above average, and for 5 subjects both resistance level and EDR frequency were below average.
Strong (up to 0.8) correlation is observed between average conductivity level and temperature.
Correlation between EDR frequency and temperature is significantly less.
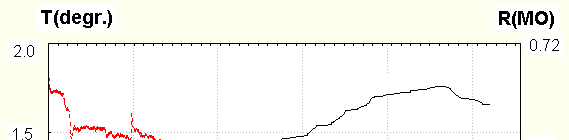
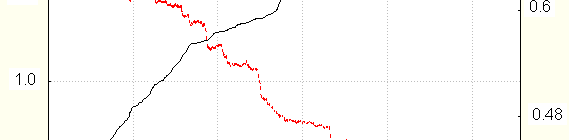
When fingers skin resistance and hand temperature were measured simultaneously, significant
crosscorrelation between them was often found : the periods of temperature increase and decrease coincide with those of resistance. Crosscorrelation coefficients could be as high as 0.8.The example is seen in Fig. 3
Resistance is solid line, left scale, temperature –dotted one, right scale.
Correlation is maximal if during resistance changes several EDR are observed. It is absent if only tonic
resistance changes are seen. In particular, it disappears during relaxation with EDR disappearance, and reappears with their beginning after activation. That is why the said correlation is found less frequently in experiments of
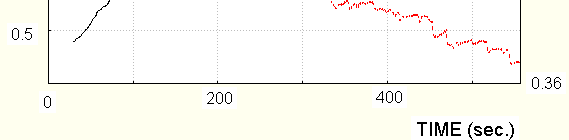
Resistance – solid line, temperature –dotted.
Correlation is significantly less, especially when - drowsiness and sleep are present (500-1000 and
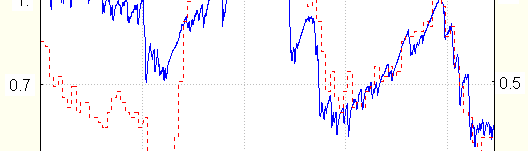
If EDA activation is obtained by transference of the subject to the room with higher temperature, (up to
27 degrees) , the resistance decreases (tonically), while the temperature increases, as should be expected for thermoregulation (Hensel 1981), that is resistance and temperature change in opposite directions. Then the temperature and conductance increase stops, and numerous EDR could be seen, see Fig.5.
Opposite trends of temperature and resistance are also observed for persons that systematically take the
medicines blocking the adrenoreceptors activity (dihydropyirydin derivatives- nifedipine and the like). It had been shown that when certain blockators concentration is achieved, the usual stimuli activating sympathetic nervous system evoke hands temperature increase instead of its usual decrease (Gulyaev 1994). In conditions mentioned, both stimulated and spontaneous EDR are absent, and stimuli cause tonic resistance decrease (seen no less than 20 times).
example, for three subjects resistance level approached maximal one for all experiments, and EDR frequency was above average, and for 5 subjects both resistance level and EDR frequency were below average.
Strong (up to 0.8) correlation is observed between average conductivity level and temperature.
Correlation between EDR frequency and temperature is significantly less.
When fingers skin resistance and hand temperature were measured simultaneously, significant
crosscorrelation between them was often found : the periods of temperature increase and decrease coincide with those of resistance. Crosscorrelation coefficients could be as high as 0.8.The example is seen in Fig. 3
Resistance is solid line, left scale, temperature –dotted one, right scale.
Correlation is maximal if during resistance changes several EDR are observed. It is absent if only tonic
resistance changes are seen. In particular, it disappears during relaxation with EDR disappearance, and reappears with their beginning after activation. That is why the said correlation is found less frequently in experiments of
Resistance – solid line, temperature –dotted.
Correlation is significantly less, especially when - drowsiness and sleep are present (500-1000 and
If EDA activation is obtained by transference of the subject to the room with higher temperature, (up to
27 degrees) , the resistance decreases (tonically), while the temperature increases, as should be expected for thermoregulation (Hensel 1981), that is resistance and temperature change in opposite directions. Then the temperature and conductance increase stops, and numerous EDR could be seen, see Fig.5.
Opposite trends of temperature and resistance are also observed for persons that systematically take the
medicines blocking the adrenoreceptors activity (dihydropyirydin derivatives- nifedipine and the like). It had been shown that when certain blockators concentration is achieved, the usual stimuli activating sympathetic nervous system evoke hands temperature increase instead of its usual decrease (Gulyaev 1994). In conditions mentioned, both stimulated and spontaneous EDR are absent, and stimuli cause tonic resistance decrease (seen no less than 20 times).