Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
20722.qxd
Effects of polysulfated glycosaminoglycan and hyaluronan on prostaglandin E2 production by cultured equine synoviocytes
Stephen P. Frean, BVSc, PhD, and Peter Lees, PhD
ly through inhibition of PGE2 production,9,11 although
Objective—To investigate effects of the anti-arthritic
glucocorticoids possess other actions, such as inhibi-
agents hyaluronan and polysulfated glycosaminogly-
tion of induction of inducible nitric oxide synthase12,13
can (PSGAG) on inflammatory metabolism in cultured
and the inducible isoform of cyclooxygenase
(ie, cyclooxygenase 2 [COX-2]).14,15 Sample Population—Synoviocytes cultured from
Pharmacologic intervention also may be directed
samples obtained from the metacarpophalangeal
at limiting damage to, and stimulating repair of, artic-
ular cartilage. To this end, glycosaminoglycan prepara-
Procedure—Equine synoviocytes were grown in
tions such as polysulfated aminoglycan (PSGAG) and
monolayer culture. Synoviocytes were stimulated
hyaluronan are administered intra-articularly or sys-
with lipopolysaccharide (LPS) and simultaneously
temically (IV administration in the case of hyaluronan
treated with various concentrations of hyaluronan orPSGAG for 48 hours. Three hyaluronan preparations
and IM administration in the case of PSGAG).
were compared. Prostaglandin E2 (PGE2) concentra-
Although these compounds can directly stimulate
tions in culture medium were measured, using
neosynthesis of cartilage matrix,16,b it is possible that
their chondroprotective effects may result indirectly
Results—The highest concentrations of hyaluronan
from a diverse range of anti-inflammatory properties.
and PSGAG tested inhibited PGE2 production.
Included in these putative anti-inflammatory actions is
Conclusions and Clinical Relevance—Clinically
achievable concentrations of hyaluronan and PSGAG
Cultured equine synoviocytes synthesize large
inhibited PGE2 synthesis by cultured equine synovio-
amounts of PGE2 in vitro when treated with
cytes. This anti-inflammatory action may be a mech-
lipopolysaccharide (LPS).21 Using the in vitro system of
anism through which these agents exert anti-arthritic
that study, we investigated the effects of various concen-
effects. The effect was obtained at concentrations
trations of hyaluronan and PSGAG on PGE2 production
that can be achieved by use of intra-articular, but not
by cultured equine synoviocytes. In light of claims by
systemic, administration of hyaluronan or PSGAG.
pharmaceutic companies of benefit of 1 hyaluronan
preparation, compared with other preparations,d,e it wasof interest to evaluate a therapeutic Streptococcus zooepi-Joint disease in horses commonly is characterized by demicus-derived hyaluronan preparation and a generic
inflammation of the synovium, producing the clas-
laboratory-grade S zooepidemicus-derived preparation.
sic signs of swelling (joint effusion), heat, and pain,1,a
Furthermore, claims have been made of greater thera-
causing loss of normal function and manifesting as
peutic effectiveness of bacterial-derived hyaluronan,
lameness. Although synovitis often is characterized by
compared with rooster comb-derived hyaluronan22;
leukocytic infiltration of the synovial membrane,2 syn-
thus, we believed it pertinent to evaluate a preparation
oviocytes can produce pro-inflammatory cytokines,3,4
of hyaluronan from this latter source.
metalloproteinases,5 and eicosanoids.6-8 Of the latter,
The purpose of the study reported here was to
prostaglandin E
investigate possible anti-inflammatory properties of
2 (PGE2) has been suggested as an
important mediator of inflammation and hyperalgesia
hyaluronan and PSGAG, which are used in the treat-
through its enhancement of vascular permeability,
ment of equine joint disease. Furthermore, the study
vasodilatory properties, and sensitization of joint noci-
was intended to reveal information about their mecha-
ceptors.9,10 Pharmacologic treatment of joint disease
nism of action, which remains poorly defined.
typically is directed at alleviating the signs of joint inflammation; nonsteroidal anti-inflammatory drugs Materials and Methods (NSAID) and synthetic glucocorticoids are the most Tissue specimens—Synovial membrane was obtained
commonly used compounds. These drugs act primari-
from the cranial and palmar joint recesses of metacarpopha-langeal (fetlock) joints of euthanatized horses. Each experi-ment (1 therapeutic agent/experiment) used pooled tissue
specimens obtained from both metacarpophalangeal joints of
the same horse. Inter-individual variation was avoided as
From the Department of Veterinary Basic Sciences, Royal Veterinary
much as possible; tissue was obtained from horses < 10 years
College, Hawkshead Ln, North Mymms, Herts AL9 7TA, UK.
old (as determined on the basis of examination of dentition).
Presented in part at the 42nd Annual Meeting of the Orthopaedic
Synovium was only used when examination of the joints, lig-
Research Society, Atlanta, Ga, Feb 22, 1996.
Support provided by the Home of Rest for Horses, Speen, UK, and
aments, and tendons in the distal aspect of each limb did not
Bayer Animal Health, Bury St Edmunds, UK.
reveal macroscopic signs of acute or chronic musculoskeletal
disease. Criteria for selection were as follows: synovial mem-
the following treatment conditions: unstimulated (medium
brane did not have signs of acute or chronic inflammation;
only), 10 µg of LPS/ml, l0 µg of LPS/ml and 10–6M
articular cartilage did not have evidence of erosion or wear
indomethacin, 10 µg of LPS/ml and various concentrations of
lines; periarticular osteophytes were not observed; synovial
PSGAG (2, 20, 200, 2,000, 10,000 and 20,000 µg/ml), PSGAG
fluid appeared normal in quantity, viscosity, and color
only (20,000 µg/ml), and 10 µg of LPS/ml and a volume of
(ie, blood was not evident); and intra- or periarticular frac-
PBS solution equivalent to that of 20,000 µg of PSGAG/ml. Prostaglandin E2 assay—Radioimmunoassay for deter- Cell culture—Synovial membrane was dissected from
mination of PGE2 concentration was conducted in accordance
the underlying adipose subintima, transferred to a Dulbecco
with the general methods described by Salmon,23 using com-
modified Eagle mediumf containing 10% (vol:vol) heat-inac-
mercially available antiserumv raised against PGE2-bovine
tivated fetal calf serumg and 1% (vol:vol) 5,000 U of peni-
serum albumin and tritiated PGE2 tracer.w Absolute PGE2
concentrations of specimens were calculated by comparison
subjected to enzymatic digestion (type VIII collagenasei from
with PGE2 standardsx of known concentration. Specimens
Clostridium histolyticum, 300 U/ml, for 3 hours at 37 C).
were diluted to ensure that values could be extrapolated from
Newly released synoviocytes were seeded in 25-cm3 flasks.j
the standard curve and were subsequently corrected for the
After incubation for 48 hours, medium was changed, remov-
original dilution. Upper and lower limits of detection of the
ing any cells not adhering to the culture flask. Flasks were
assay were 10 ng/ml and 50 pg/ml, respectively.
maintained by changing medium at 48-hour intervals until
Statistical analysis—Statistical analysis was conducted,
cell replication was observed; at that time, synoviocytes were
using a proprietary statistical software program.y For each
seeded in 24-well platesk and grown to confluence.
therapeutic agent, results represented the data of 1 experi-
Procedures involving hyaluronan—The dose of
ment, with 4 replicates in each treatment group (ie, n = 4).
hyaluronan typically recommended for intra-articular admin-
Values were expressed as mean ± SEM. Significant differences
istration in horses is 20 mg/joint.l Typically, 5 to 6 ml of syn-
in PGE2 synthesis in response to various treatments were
ovial fluid can be aspirated from a fetlock joint of a clinical-
determined by use of one-way ANOVA followed by use of the
ly normal horse. Therefore, immediately subsequent to intra-
Dunnett test. Significance was assigned at P < 0.05.
articular injection, it is possible to achieve a concentration of3 to 4 mg of hyaluronan/ml. An inflamed joint typically will
be swollen as a result of synovial effusion; administration of
Commercially available hyaluronan prepara-
20 mg of hyaluronan will result in concentrations in the syn-
tion—Synoviocytes incubated with 10 µg of LPS/ml
ovial space that are < 4 mg/ml. The rate of decrease in con-
produced concentrations of PGE2 significantly
centration after intra-articular administration is not known;
(P < 0.001) higher than the basal concentrations of
therefore, hyaluronan was used at concentrations of 20, 200,
unstimulated cells. The LPS-indomethacin positive-
500, 1,000 and 2,000 µg/ml. Because addition of the highest
control treatment caused a significantly (P < 0.001)
concentration of hyaluronan (ie, 2,000 µg/ml) resulted in
dilution of culture medium to 80% of normal concentration,
control treatments were incorporated into the experiment. Volumes of phosphate-buffered saline (PBS) solutionm equivalent to volumes for the highest concentrations of hyaluronan (ie, 1,000 and 2,000 µg/ml) were added with LPSn (10 µg/ml) to culture wells. Indomethacin, a NSAID that inhibits PGE2 synthesis, was used as a positive-control
Synoviocytes were incubated for 48 hours, using the fol-
lowing treatment conditions: unstimulated (medium only),10 µg of LPS (Escherichia coli serotype B5:055)/ml, 10 µg ofLPS/ml and 10–6M indomethacin,o 10 µg of LPS/ml and vari-ous concentrations of hyaluronan (ie, 20, 200, 500, 1,000and 2,000 µg/ml), hyaluronan only (1,000 and 2,000 µg/ml),and 10 µg of LPS/ml and PBS solution equivalent in volumeto the volumes of 1,000 and 2,000 µg of hyaluronan/ml.
In addition to a commercially available preparation of
hyaluronan,p generic preparations of hyaluronan formulatedfrom S zooepidemicusq and rooster combr were tested. Theselatter hyaluronan preparations were obtained in solid form. To simulate as closely as possible the preparation of the com-mercially available product, the generic products were dis-solved in PBS solution (initial concentration, 10,000 µg/ml).
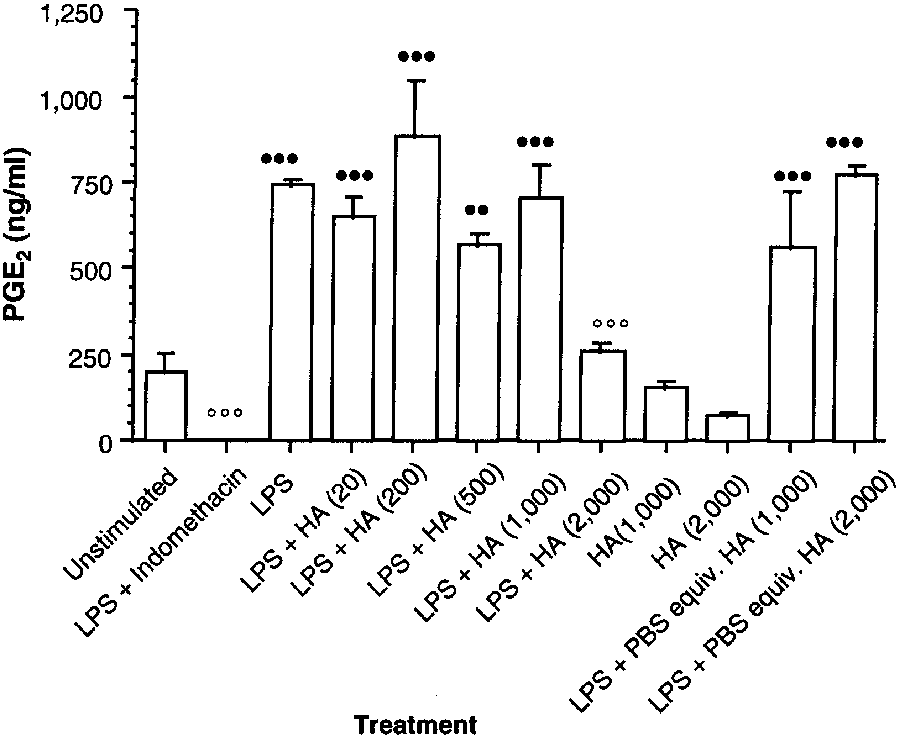
Figure 1—Effect of a commercially available hyaluronan (HA)preparation on prostaglandin E2 (PGE2) synthesis by lipopolysac-
Procedures involving polysulfated glycosamino-
charide (LPS)-stimulated synoviocytes obtained from the
glycan—At the clinically recommended dose for horses of
metacarpophalangeal joints of clinically normal horses. Synoviocytes were incubated for 48 hours at 37 C. Each treat-
250 mg of PSGAG/joint,s the highest concentration achievable
ment group represents mean ± SEM PGE2 concentration of 4
by intra-articular injection of a fetlock joint is approximately
replicate wells. Hyaluronan concentrations (in parentheses) are
50 mg/ml. The recommended dose for IM administration of
expressed in µg/ml. The LPS concentration is 10 µg/ml.
PSGAG in horses is 500 mg/horse.t A theoretical maximum
Indomethacin concentration is 10-6M. Values differ significantly
plasma concentration of 0.025 mg/ml can be calculated for
(n = P < 0.05, nn = P < 0.01, nnn = P < 0.001) from those forunstimulated synoviocytes (Dunnett test, 2-tailed). Values differ
horses, assuming rapid and complete absorption from the
significantly (o = P < 0.05, oo = P < 0.01, ooo = P < 0.001) from
injection site. Therefore, various concentrations of PSGAGu
those for synoviocytes incubated with LPS (Dunnett test,
were tested. Synoviocytes were incubated for 48 hours, using
2-tailed). PBS = Phosphate-buffered saline solution.
Table 1—Effect of various concentrations of 3 hyaluronan prepa-rations on the percentage change in prostaglandin E2 synthesis
by lipopolysaccharide (LPS)-stimulated synoviocytes obtainedfrom the metacarpophalangeal joints of clinically normal horses
Hyaluronan preparation Commercially Streptococcus Treatment* available zooepidemicus
Positive values reflect increased PGE2 synthesis, negative values
reflect decreased PGE2 synthesis, compared with values for synovio-
*Concentration of LPS = 10 g/ml, and hyaluronan concentrations
a,bValues are significantly (a, P Ͻ 0.01; b, P Ͻ 0.001) different from
Figure 2—Effect of a Streptococcus zooepidemicus HA prepa-
those for synoviocytes incubated in LPS alone (Dunnett test, 2-tailed).
ration on PGE2 synthesis by LPS-stimulated synoviocytes
obtained from the metacarpophalangeal joints of clinically nor-mal horses. See Figure 1 for key. Streptococcus zooepidemicus hyaluronan prepa- ration—Incubation of ynoviocytes with LPS signifi- cantly (P < 0.001) increased PGE2 concentrations,
compared with values for basal concentrations ofunstimulated cells. The LPS-indomethacin positive-control treatment significantly (P < 0.001) reducedPGE2 synthesis, compared with values for LPS alone.
Synoviocytes incubated with hyaluronan (1,000 and2,000 µg/ml) had slight, but not significant, decreas-es in PGE2 synthesis of 8 and 11%, respectively.
Incubation of LPS-stimulated cultures with PBS solu-tion equivalent in volume to 1,000 and 2,000 µg ofhyaluronan/ml significantly (P < 0.001) increasedPGE2 synthesis, compared with values for unstimu-
lated cells; value for the cells incubated with LPS- PBS solution was similar to that of cells treated with LPS alone (Fig 2). Similar to the commercially avail- able hyaluronan preparation, the S zooepidemicus hyaluronan preparation at low concentrations did
Figure 3—Effect of a rooster comb HA preparation on PGE2 syn-
not cause significant effects on LPS-enhanced PGE2
thesis by LPS-stimulated synoviocytes obtained from the
synthesis (Table 1). Similar to the commercially
metacarpophalangeal joints of clinically normal horses. See
available hyaluronan preparation, however, the high-
est concentration of S zooepidemicus hyaluronanpreparation (2,000 µg/ml) produced a significant
with synoviocytes incubated with 10 µg of LPS/ml.
(P = 0.007) reduction (56%) in PGE
Unstimulated synoviocytes incubated with the com-
pared with that for cells incubated with LPS alone.
mercially available preparation at concentrations of1,000 and 2,000 µg/ml caused decreases in PGE2 syn-
Rooster-comb hyaluronan preparation—Unstim-
thesis of 21 and 61%, respectively, but these values were
ulated synoviocytes had a higher basal PGE2 synthesis
not significantly different (Fig 1). The LPS-stimulated
than unstimulated synoviocytes in the other 2 experi-
cultures incubated with PBS solution equivalent in vol-
ments involving the use of hyaluronan. Incubation with
ume to 1,000 and 2,000 µg of hyaluronan/ml had sig-
LPS produced a comparatively weaker, nonsignificant
nificantly (P < 0.001) increased PGE2 synthesis, com-
increase in PGE2 concentration, although mean absolute
pared with values for unstimulated cells. The LPS-stim-
PGE2 concentration (724 ng/ml) was similar to that pro-
ulated cells treated with hyaluronan (20, 200, 500 and
duced by use of LPS in the other 2 experiments involv-
1,000 µg/ml) did not have significant changes in the
ing hyaluronan (Fig 3). Incubation of LPS-stimulated
enhanced PGE2 synthesis, compared with values for
cultures with PBS solution equivalent in volume to
LPS-stimulated cells not treated with hyaluronan
hyaluronan (1,000 and 2,000 µg/ml) enhanced PGE2
(Table 1). The highest concentration of the commer-
synthesis similar to that of cells treated with LPS alone.
cially available hyaluronan preparation (2,000 µg/ml)
In contrast to the generic S zooepidemicus and commer-
caused a significant (P < 0.001) decrease in PGE2 pro-
cially available hyaluronan preparation, rooster comb
duction, compared with PGE2 concentrations for cells
hyaluronan (1,000 µg/ml) enhanced synthesis of PGE2
(increase of 83%) in unstimulated cells, which was not
oviocytes; macrophage-like synovial cells cultured fromsynovium obtained from osteoarthritic humans had adecrease in interleukin-1α-induced PGE2 production
when treated with hyaluronan at concentrations of 0.5and 1 mg/ml.20 This decrease in PGE2 synthesis also was
dependent on hyaluronan molecular weight. Lapinechondrocytes stimulated with interleukin-1 similarlyhad a decrease in PGE2 synthesis when treated with
hyaluronan at similar concentrations to those used onhuman cells17; in that study, hyaluronan had a concen-tration- (0.5, 1 and 2 mg/ml) and molecular weight-dependent (0.5, 1, and 2 kd) inhibition of PGE2 syn-
thesis. The exact molecular weight of each hyaluronanpreparation used in the study reported here is notknown. However, the commercially available hyaluro-nan preparation used in our study is of lower viscositythan other commercially available hyaluronan prepara-tions licensed for intra-articular administration.22
Analysis of the data in the study reported here failed
to confirm the findings of other investigators.17,20 None ofthe 3 hyaluronan preparations, at concentrations up to
Figure 4—Effect of polysulfated glycosaminoglycan (PSGAG) on
1,000 µg/ml, exerted inhibitory actions on LPS-induced
PGE2 synthesis by LPS-stimulated synoviocytes obtained from
PGE2 synthesis. However, at the highest concentration
the metacarpophalangeal joints of clinically normal horses. Eachtreatment group represents mean ± SEM PGE
(2,000 µg/ml), 2 of the 3 preparations (commercially
4 replicate wells. The PSGAG concentrations (in parentheses)
available preparation and S zooepidemicus preparation)
are expressed in µg/ml. See Figure 1 for key.
significantly inhibited but, unlike indomethacin, did notabolish PGE
significant (P = 0.08). This value compared favorably
2 production. The reason for the failure of
the rooster comb hyaluronan preparation to inhibit PGE
with the slight (7%) increase produced by 2,000 µg of
synthesis at each concentration is not known. It may
hyaluronan/ml, which also was not significant.
have been attributable to a low molecular weight of that
Treatment of LPS-stimulated cells with rooster comb
preparation. Indeed, low concentrations of this prepara-
hyaluronan increased production of PGE2; this
tion failed to inhibit and actually increased PGE
enhanced response was somewhat greater at lower con-
sis beyond that caused by LPS alone.
centrations of hyaluronan (20 µg/ml = 76%, P < 0.001;
A possible explanation for the increased PGE
200 µg/ml = 49%; 500 µg/ml = 40%; 1,000 µg/ml = 19%;
concentrations measured in cultures treated with
rooster comb hyaluronan is that there may be pro-
Polysulfated glycosaminoglycan preparation—
inflammatory contaminants in the preparation, as has
Incubation of synoviocytes with LPS caused PGE
been suggested elsewhere.22 On the basis of this suppo-
thesis that was significantly (P = 0.006) greater than the
sition, it could be suggested that the enhanced PGE2
basal concentration of unstimulated cells. The LPS-
synthesis obtained in our study may have been coun-
indomethacin positive-control treatment significantly
tered only by higher concentrations of hyaluronan
ues for LPS alone. Incubation of synoviocytes with
potential pro-inflammatory contaminants. However, it
PSGAG (20,000 µg/ml) also profoundly decreased (82%)
would appear that enhanced PGE2 synthesis was not
an artifact resulting from contaminated PBS solvent,
2 synthesis. The LPS-stimulated cells treated with 2
µg of PSGAG/ml were unaffected, but treatment with 20
because the same stock of PBS solution was used
µg of PSGAG/ml caused a significant (P = 0.035) increase
throughout the study. Furthermore, LPS-stimulated
of 44% above the concentrations for cells treated with
cells incubated with a volume of PBS solution equiva-
LPS alone (Fig 4). In contrast, 4 higher concentrations of
lent to that for hyaluronan concentrations of 1,000 and
PSGAG (200, 2,000, 10,000, and 20,000 µg/ml)
2,000 µg/ml had similar PGE2 synthesis to those cells
(decreases of 32, 58, 99, and 96%, respectively); howev-
The reduction in PGE2 production in response to
er, only the latter 2 decreases were significantly
2 hyaluronan preparations (commercially available and
(P = 0.003) different from the value for cells stimulated
S zooepidemicus preparations) at the highest concentra-
by LPS alone. The control treatment (ie, incubation of
tions used (2,000 µg/ml) as a result of mere dilution of
cells with LPS and a volume of PBS solution equivalent
the culture medium to 80% of its original concentra-
to that for 20 µg of PSGAG/ml) produced a value for
tion can be discounted. In control cells treated with
LPS in medium diluted with a volume of PBS solution
2 synthesis similar to that of LPS alone.
equivalent to that for hyaluronan at 1,000 and 2,000 µg/ml, PGE2 concentrations were similar to
Discussion
those obtained in LPS-stimulated cultures maintained
Other investigators have documented that hyaluro-
nan inhibits PGE2 synthesis by cultured human syn-
Using unstimulated hamster kidney cells, Dietmar18
reported that PSGAG at concentrations of 0.03, 0.1, 0.3,
hyaluronan inhibits the release of PGE2 into culture
and 1% reduced PGE2 concentrations in culture super-
medium, rather than inhibiting the synthesis of PGE2,
natant in a concentration-dependent manner at 16
is worthy of consideration. Some investigators have
hours (44 to 52%), and 48 hours (35 to 67%) after ini-
suggested that the diverse actions of hyaluronan on
tiation of incubation. The PSGAG used in that study is
cellular metabolism may be a consequence of its high
a preparation licensed for use in humans; that prepara-
viscosity in solution; it has been suggested that
tion, similar to the preparation licensed for use in hors-
hyaluronan could physically retard the release of PGE2
es, contains 250 mg of PSGAG/ml. Conversion of units
from cells, as opposed to inhibiting synthesis.17,20
of concentration used in our study (µg/ml) to those
Similarly, it has been suggested that hyaluronan may
used by Dietmar (percentage) indicates that reductions
prevent mobilization of eicosanoid precursors. Release
in LPS-induced PGE2 synthesis in cultures of equine
of 14C-labeled arachidonic acid from human synovial
synoviocytes were obtained at PSGAG concentrations
fibroblasts in response to bradykinin and calcium
of 0.02, 0.2, 1, and 2%, similar to those used with ham-
ionophore is inhibited by hyaluronan in both a molec-
ster kidney cells. The lower concentrations used in the
ular weight- and concentration-dependent manner.24
study reported here (ie, 0.002 and 0.0002%) did not
However, authors of that study concluded that inhibi-
tion of arachidonic acid release was not associated with
We were unable to account for the increase above
viscosity, because methylcellulose, a liquid similarly
baseline values of PGE2 synthesis by LPS-stimulated cells
viscous to hyaluronan, did not inhibit arachidonic acid
treated with 20 µg of PSGAG/ml. Results of experiments
had little intra-experiment variation, and conditions
Binding interactions of polysaccharide prepara-
were standardized with regard to number of cells and
tions, such as pentosan polysulfate and hyaluronan,
population. Therefore, it is unlikely that these potential-
with cell membrane molecules have received attention.
ly variable factors accounted for the increase in PGE2
Thrombospondin has been suggested as a receptor
concentration. Thus, the implication is that the increase
through which pentosan polysulfate may act as a
in this treatment group may have been the result of a pos-
matrix metalloproteinase inhibitor.25 Cellular receptors
sible synergistic effect of LPS and PSGAG on PGE2 syn-
for hyaluronan have been identified, and it has been
thesis at that concentration of the compound. However,
suggested that adhesion and migration of cells may be
this is unlikely in view of the remainder of the data.
modulated through these hyaluronan receptors.26-29
Similar to our observations of decreased PGE2 syn-
Also, the capacity of hyaluronan to form complexes
thesis in response to hyaluronan, decreased PGE2 con-
with phospholipids has been described recently,30,31,z,aa
centrations were obtained in cultures treated with con-
with the suggestion that administration of hyaluronan
centrations of PSGAG similar to those estimated to be
into the joint space may provide a source of binding
obtainable by intra-articular injection, but not at con-
sites for phospholipids and their metabolites (found in
centrations that can be achieved after IM administration.
appreciable amounts in synovial fluid and synovial tis-
Synoviocytes for each experiment were cultured
sues of arthritic joints), which may stimulate chondro-
from tissue of specific horses, which varied among
cyte catabolism. Consequently, hyaluronan may exert a
experiments. All other experimental conditions were
chondroprotective effect by binding these pro-inflam-
standardized, and batches of reagents and equipment
matory (or pro-chondrodestructive) molecules. The
were common to all experiments. Therefore, it is like-
observation in the study reported here of reduced PGE2
ly that the differences in absolute baseline and LPS-
synthesis after incubation with hyaluronan may be
stimulated PGE2 concentrations measured in each
explained by the binding capacity of hyaluronan, on
experiment resulted from variation among horses,
the basis that the production of eicosanoids such as
despite efforts to standardize selection of synovium
PGE2 results from metabolism of cell membrane phos-
from nonarthritic joints in horses of similar age.
pholipids. Hyaluronan may bind these phospholipids
In the study reported here, we measured reduc-
in stable complexes, rendering them less susceptible to
tions in the concentration of PGE2 in culture medium
breakdown. This hypothesis has not been investigated.
obtained from monolayer cultures of equine synovial
Using a model of osteoarthritis in which a carpal
cells. However, these experiments did not address the
chip was created surgically, Kawcak et al2 measured
exact mechanism of action of hyaluronan and PSGAG
PGE2 and protein concentrations in synovial fluid of
at the cellular and molecular levels. Exposure of many
horses treated by IV administration of hyaluronan.
cell types to LPS results in induction of COX-2, with
Compared with concentrations for untreated control
consequent synthesis of PGE2 by that inducible iso-
horses, synovial fluid concentrations of PGE2 and pro-
form of cyclooxygenase. Mechanisms of action of
tein were decreased in hyaluronan-treated joints.
NSAID and corticosteroids in inhibiting inflammatory
Furthermore, histologic examination of the synovial
eicosanoid synthesis are established (inhibition of the
membranes of surgically explored joints revealed
actions of COX-2 and phospholipase A2 respectively,
decreased synovial vascularity and cellular infiltration
and, in the case of corticosteroids, inhibition of induc-
in horses treated by use of IV administration of
tion of the COX-2 isoenzyme as well). Given the dis-
hyaluronan, compared with surgically explored joints
similarity in molecular structure of hyaluronan and
of untreated control horses.2 The authors of that study
PSGAG to corticosteroids and NSAID, it may be con-
concluded that IV administration of hyaluronan had a
sidered unlikely that hyaluronan and PSGAG exert
positive effect in ameliorating inflammatory synovitis,
their PGE2 concentration-lowering effects through
although the mechanism of action of hyaluronan
inhibition of these enzymes. The possibility that
remains unknown. The data from our in vitro study
warrant discussion because of results of the aforemen-
induces receptor shredding from synovial leukocytes (abstr). Br J
tioned in vivo study. Our calculations of the plasma
Rheumatol 1995;34(suppl 34):30.
and synovial fluid concentrations of hyaluronan thatmay theoretically be obtained following IV injection
References
suggest that the decreased synovial fluid concentra-
1. Owens JG, Kamerling SG, Stanton SR, et al. Effects of pre-
treatment with ketoprofen and phenylbutazone on experimentally
2 observed by Kawcak et al2 were obtained
in response to concentrations of hyaluronan far less
induced synovitis in horses. Am J Vet Res 1996;57:866–874.
2. Kawcak CE, Frisbie DD, Trotter GW, et al. Effects of intra-
than those (1,000 to 2,000 µg/ml) required to decrease
venous administration of sodium hyaluronate on carpal joints in
PGE2 concentrations in culture medium in our experi-
exercising horses after arthroscopic surgery and osteochondral frag-
ments. Although equine synovial fibroblasts are a
mentation. Am J Vet Res 1997;58:1132–1140.
3. Dingle JT, Saklatvala J, Hembry R, et al. A cartilage catabol-
infiltrating the synovial membrane in horses with syn-
ic factor from synovium. Biochem J 1979;184:177–180.
ovitis are another important source of PGE
4. Saklatvala J, Dingle JT. Identification of catabolin, a protein
from synovium which induces the degradation of cartilage in organ
pro-inflammatory mediators. Therefore, it may be that
culture. Biochem Biophys Res Commun 1980;96:1225–1231.
the decreased synovial fluid concentrations of PGE2
5. Okada Y, Shinmei M, Tanaka O, et al. Localisation of matrix
reported by Kawcak et al2 are a consequence of
metalloproteinase 3 (stromelysin) in osteoarthritic cartilage and syn-
hyaluronan’s effects on leukocytes that infiltrated the
ovium. Lab Invest 1994;66:680–690.
synovium. On the basis of reported inhibitory effects of
6. Hulkower KI, Coffey JW, Levin W, et al. Interleukin-1b
hyaluronan on leukocyte migration32,33 and the identifi-
induces cytosolic PLA2 in parallel with prostaglandin E2 in rheuma-toid synovial fibroblasts. Agents Actions 1993;39:C5–C7.
cation of hyaluronan receptors on leukocytes,34,bb it may
7. Lindsley HB, Smith DD. Enhanced prostaglandin E2 secre-
be that the anti-inflammatory effects of hyaluronan in
tion by cytokine-stimulated human synoviocytes in the presence of
that in vivo experimental model of equine joint disease
subtherapeutic concentrations of nonsteroidal antiinflammatory
reside in the drug’s actions on leukocytes as well as on
drugs. Arthritis Rheum 1990;33:1162–1169.
the synovial membrane. In both instances, it would be
8. May S, Hooke R, Lees P. Identity of the E-series
necessary to assume that hyaluronan can exert its
prostaglandin produced by equine chondrocytes and synovial cells inresponse to a variety of stimuli. Res Vet Sci 1989;46:54–57.
actions without penetrating the membrane, because
9. Palmer J, Bertone A. Joint structure, biochemistry and bio-
this is unlikely, given its high molecular weight.
chemical disequilibrium in synovitis and equine joint disease. Equine Vet J 1994;26:263–277.
aPalmer JL, Bertone AL. Effect of experimentally-induced acute syn-
10. May SA, Hooke RE, Perelmans KY, et al. Prostaglandin E2 in
ovitis on articular cartilage proteoglycan synthesis and composi-
equine joint disease. Vlaams Diergeneeskd Tijdschr 1994;63:
tion (abstr). Trans Orthop Res Soc 1994;19:480.
bFrondoza C, Chang D, Jeng C, et al. Hyaluronan enhances proteo-
11. Lees P, May SA, Cambridge H, et al. Pathophysiology of
glycan synthesis of human chondrocytes and synoviocytes (abstr).
inflammation and joint disease mechanisms in the horse: mecha-
Osteoarthritis Cartilage 1994;2(suppl 1):48.
nisms, mediators and medicines, in Proceedings. 4th Congr Vet Assoc
cTodesco S, Punzi L. Influence of intra-articular injection of
Vet Pharmacol Toxicol 1988;197–204.
hyaluronic acid (HA) on synovial fluid (SF) inflammation (abstr).
12. Simmons WW, Ungureanu-Longrois D, Smith GK, et al. Osteoarthritis Cartilage 1993;1:69.
Glucocorticoids regulate inducible nitric oxide synthase by inhibit-
dProduct Information Monograph, Bayer AG, Leverkusen, Germany.
ing tetrahydrobiopterin synthesis and L-arginine transport. J Biol
eProduct Information Monograph, Pharmacia AB, Stockholm, Sweden.
fDulbecco’s Modified Eagle’s Medium, Life Technologies Ltd,
13. Kleinert H, Euchenhofer C, Ihrig-Biedert I, et al.
Glucocorticoids inhibit the induction of nitric oxide synthase II by
gFetal Calf Serum, Harlan Sera-Lab Ltd, Loughborough, UK.
down-regulating cytokine-induced activity of transcription factor
hPenicillin/Streptomycin Solution, Life Technologies Ltd, Paisley, UK.
nuclear factor-kappa B. Mol Pharmacol 1996;49:15–21.
iCollagenase, Sigma Aldrich Co, Poole, UK.
14. Morisset S, Patry C, Lora M, et al. Regulation of cyclooxy-
jTissue Culture Flask 25-cm3, Life Technologies Ltd, Paisley, UK.
genase-2 expression in bovine chondrocytes in culture by interleukin
kTissue Culture Plate 24-well, Life Technologies Ltd, Paisley, UK.
1 alpha, tumor necrosis factor-alpha, glucocorticoids, and 17beta-
lProduct Information Monograph, Bayer AG, Leverkusen, Germany.
estradiol. J Rheumatol 1998;25:1146–1153.
mPhosphate buffered saline, Life Technologies Ltd, Paisley, UK.
15. Newton R, Kuitert LM, Slater DM, et al. Cytokine induction
nLipopolysaccharide, Sigma Aldrich Co, Poole, UK.
of cytosolic phospholipase A2 and cyclooxygenase-2 mRNA is sup-
oIndomethacin, Sigma Aldrich Co, Poole, UK.
pressed by glucocorticoids in human epithelial cells. Life Sci
pHyonate, Bayer Animal Health, Bury St Edmunds, UK.
qHyaluronic Acid (from S zooepidemicus), Sigma Aldrich Co, Poole, UK.
16. Glade M. Polysulfated glycosaminoglycan accelerates net
rHyaluronic Acid (from rooster comb), Sigma Aldrich Co, Poole, UK.
synthesis of collagen and glycosaminoglycans by arthritic equine car-
sProduct Information Monograph, Luitpold Pharmaceuticals Inc,
tilage tissues and chondrocytes. Am J Vet Res 1990;51:779–785.
17. Akatsuka M, Yamamoto Y, Tobetto K, et al. In vitro effects of
tProduct Information Monograph, Luitpold Pharmaceuticals Inc,
hyaluronan on prostaglandin E2 induction by interleukin-1 in rabbit
articular chondrocytes. Agents Actions 1993;38:122–125.
uAdequan, Janssen Animal Health, High Wycombe, UK.
18. Dietmar EGG. Effects of glycosaminoglycan polysulphate
vAnti-Prostaglandin E2 Whole Antiserum, Sigma Aldrich Co,
and two non-steroidal anti-inflammatory drugs on prostaglandin E2
synthesis in Chinese hamster ovary cell cultures. Pharmacol Res
w[3H]Prostaglandin E2, Amersham Pharmacia Biotech, Little
19. Punzi L, Schiavon F, Cavasin F, et al. The influence of intra-
xProstaglandin E2, Sigma Aldrich Co, Poole, UK.
articular hyaluronic acid on PGE2 and cAMP of synovial fluid. ClinExp Rheumatol 1989;7:247–250.
zPasquali-Ronchetti I. Hyaluronan-phospholipid interactions: causal-
20. Yasui T, Akatsuka M, Tobetto K, et al. The effect of hyaluro-
ity or biological role (abstr). Immunol Cell Biol 1993;73:A21.
nan on interleukin-1α-induced prostaglandin E2 production in
aaGhosh P. Hyaluronan and biochemical lesions of osteoarthritis
human osteoarthritic synovial cells. Agents Actions 1992;37:155–156.
(abstr). Immunol Cell Biol 1996;74:A9.
21. Landoni M, Foot R, Frean S, et al. Effects of flunixin, tolfe-
bbBrennan FR, Mikecz K, Jobanputra P, et al. An anti-CD44 antibody
namic acid, R(-) and S(+) ketoprofen on the response of equine syn-
oviocytes to lipopolysaccharide stimulation. Equine Vet J 1996;28:
tors: key players in growth, differentiation, migration and tumor pro-
gression. Curr Opin Cell Biol 1994;6:726–733.
22. Brown K. Legend®, from concept to product, in Proceedings.
30. Ghosh P, Holbert C, Read R, et al. Hyaluronic acid (hyaluro-
Hyaluronic Acid Workshop, Bayer Corp, Kansas City 1995 1995:6–10.
nan) in experimental osteoarthritis. J Rheumatol 1995;22(suppl 43):
23. Salmon JA. A radioimmunoassay for 6-keto-prostaglandin
F1α. Prostaglandins 1978;15:383–397.
31. Ghosh P, Hutadilok N, Adam N. Interactions of hyaluronan
24. Tobetto K, Yasui T, Ando T, et al. Inhibitory effects of
(hyaluronic acid) with phospholipids as determined by gel perme-
hyaluronan on [14C]arachidonic acid release from labeled human
ation chromatography, multi-angle laser-light scattering photo-
synovial fibroblasts. Jpn J Pharmacol 1992;60:79–84.
metry and 1H-NMR spectroscopy. Int J Biol Macromolecules
25. Ghosh P, Hutadilok N. Interactions of pentosan polysulfate
with cartilage matrix proteins and synovial fibroblasts derived from
32. Miyazaki K, Gotoh S, Ohkawara H, et al. Studies on anal-
patients with osteoarthritis. Osteoarthritis Cartilage 1996;4:43–53.
gesic and antiinflammatory effects of sodium hyaluronate.
26. Aruffo A, Stamenkovich I, Melnick M, et al. CD44 is the prin-
Pharmacometrics 1984;28:1123–1135.
cipal cell surface receptor for hyaluronate. Cell 1990;61:1303–1313.
33. Tamoto K, Tada M, Shimada S, et al. Effects of high-molec-
27. Perschl A, Lesley J, English N, et al. Role of CD44 cytoplas-
ular-weight hyaluronates on the functions of guinea pig polymor-
mic domain in hyaluronan binding. Eur J Immunol 1995;25:495–501.
phonuclear leucocytes. Semin Arthritis Rheum 1993;22:4–8.
28. Stamenkovic I, Aruffo A. Hyaluronic acid receptors.
34. Mikecz K, Brennan FR, Kim JH, et al. Anti-CD44 treatment
Methods Enzymol 1994;245:195–216.
abrogates tissue oedema and leukocyte infiltration in murine arthritis.
29. Sherman L, Sleeman J, Herrlich P, et al. Hyaluronate recep-
Drugs for Alzheimer’s disease How and where can you get the drugs? The drugs that are currently available are not a cure and do not stop the The drugs are available on NHS prescription from progression of the disease. They may, however, temporarily ease some approved hospital specialists according to strict of the symptoms of Alzheimer’s disease in some people. criteria. Treatme
James Robert Brown University of Toronto The idea of a community of science is one we all hold dear. We think of ourselves — allacademics, not just scientists in the narrow sense — as pursuing common goals and doing so in anon-competitive way. To be sure, there are rivalries, often bitter. And no doubt we would alllike the recognition that comes with being the acknowledged discoverer
 disease. Criteria for selection were as follows: synovial mem-
the following treatment conditions: unstimulated (medium
brane did not have signs of acute or chronic inflammation;
only), 10 µg of LPS/ml, l0 µg of LPS/ml and 10–6M
articular cartilage did not have evidence of erosion or wear
indomethacin, 10 µg of LPS/ml and various concentrations of
lines; periarticular osteophytes were not observed; synovial
PSGAG (2, 20, 200, 2,000, 10,000 and 20,000 µg/ml), PSGAG
fluid appeared normal in quantity, viscosity, and color
only (20,000 µg/ml), and 10 µg of LPS/ml and a volume of
(ie, blood was not evident); and intra- or periarticular frac-
PBS solution equivalent to that of 20,000 µg of PSGAG/ml.
disease. Criteria for selection were as follows: synovial mem-
the following treatment conditions: unstimulated (medium
brane did not have signs of acute or chronic inflammation;
only), 10 µg of LPS/ml, l0 µg of LPS/ml and 10–6M
articular cartilage did not have evidence of erosion or wear
indomethacin, 10 µg of LPS/ml and various concentrations of
lines; periarticular osteophytes were not observed; synovial
PSGAG (2, 20, 200, 2,000, 10,000 and 20,000 µg/ml), PSGAG
fluid appeared normal in quantity, viscosity, and color
only (20,000 µg/ml), and 10 µg of LPS/ml and a volume of
(ie, blood was not evident); and intra- or periarticular frac-
PBS solution equivalent to that of 20,000 µg of PSGAG/ml.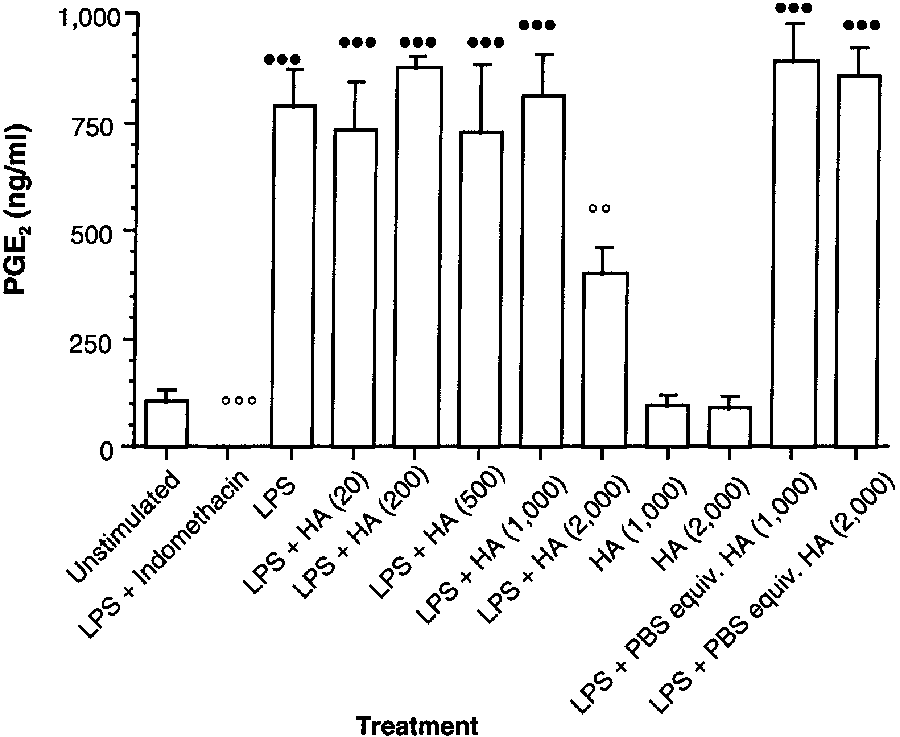
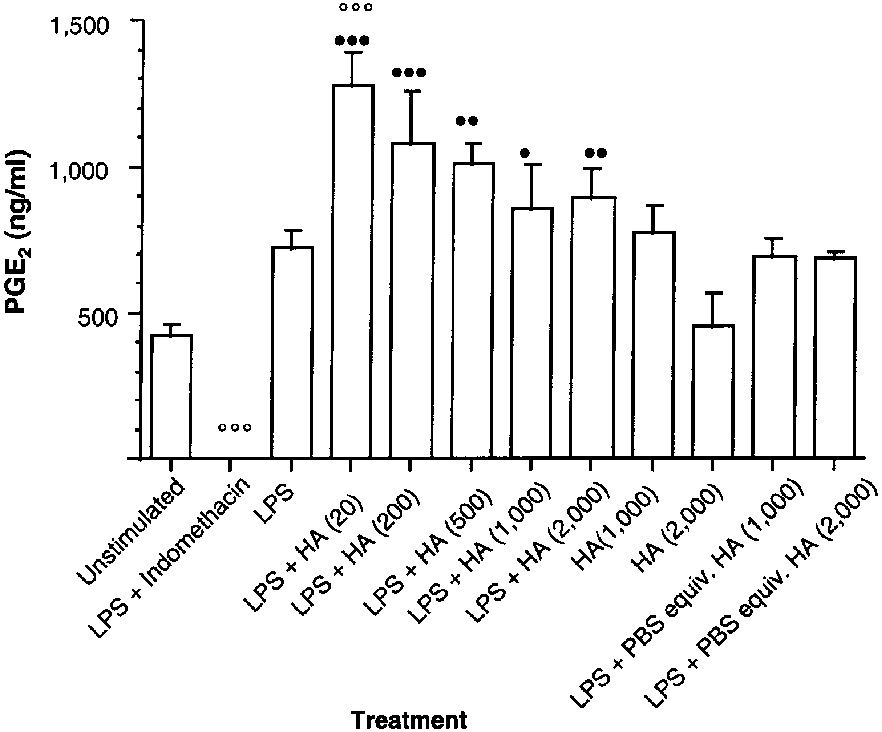 Table 1—Effect of various concentrations of 3 hyaluronan prepa-rations on the percentage change in prostaglandin E2 synthesis
by lipopolysaccharide (LPS)-stimulated synoviocytes obtainedfrom the metacarpophalangeal joints of clinically normal horses
Hyaluronan preparation
Table 1—Effect of various concentrations of 3 hyaluronan prepa-rations on the percentage change in prostaglandin E2 synthesis
by lipopolysaccharide (LPS)-stimulated synoviocytes obtainedfrom the metacarpophalangeal joints of clinically normal horses
Hyaluronan preparation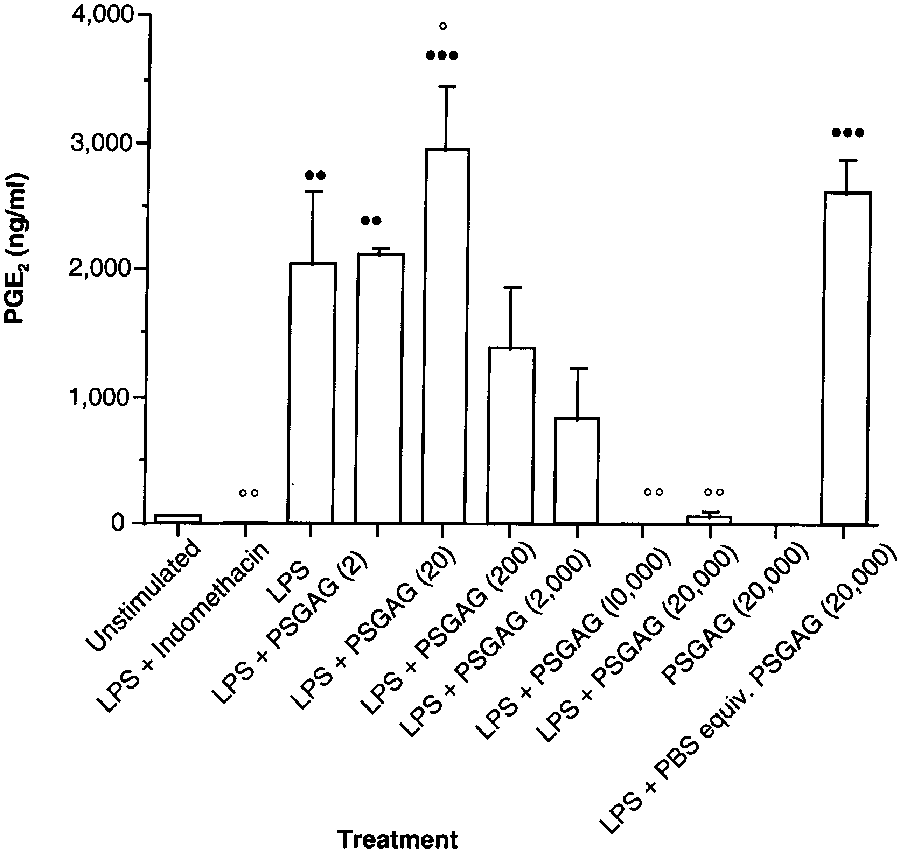 oviocytes; macrophage-like synovial cells cultured fromsynovium obtained from osteoarthritic humans had adecrease in interleukin-1α-induced PGE2 production
when treated with hyaluronan at concentrations of 0.5and 1 mg/ml.20 This decrease in PGE2 synthesis also was
dependent on hyaluronan molecular weight. Lapinechondrocytes stimulated with interleukin-1 similarlyhad a decrease in PGE2 synthesis when treated with
hyaluronan at similar concentrations to those used onhuman cells17; in that study, hyaluronan had a concen-tration- (0.5, 1 and 2 mg/ml) and molecular weight-dependent (0.5, 1, and 2 kd) inhibition of PGE2 syn-
thesis. The exact molecular weight of each hyaluronanpreparation used in the study reported here is notknown. However, the commercially available hyaluro-nan preparation used in our study is of lower viscositythan other commercially available hyaluronan prepara-tions licensed for intra-articular administration.22
Analysis of the data in the study reported here failed
to confirm the findings of other investigators.17,20 None ofthe 3 hyaluronan preparations, at concentrations up to
Figure 4—Effect of polysulfated glycosaminoglycan (PSGAG) on
1,000 µg/ml, exerted inhibitory actions on LPS-induced
PGE2 synthesis by LPS-stimulated synoviocytes obtained from
PGE2 synthesis. However, at the highest concentration
the metacarpophalangeal joints of clinically normal horses. Eachtreatment group represents mean ± SEM PGE
(2,000 µg/ml), 2 of the 3 preparations (commercially
4 replicate wells. The PSGAG concentrations (in parentheses)
available preparation and S zooepidemicus preparation)
are expressed in µg/ml. See Figure 1 for key.
oviocytes; macrophage-like synovial cells cultured fromsynovium obtained from osteoarthritic humans had adecrease in interleukin-1α-induced PGE2 production
when treated with hyaluronan at concentrations of 0.5and 1 mg/ml.20 This decrease in PGE2 synthesis also was
dependent on hyaluronan molecular weight. Lapinechondrocytes stimulated with interleukin-1 similarlyhad a decrease in PGE2 synthesis when treated with
hyaluronan at similar concentrations to those used onhuman cells17; in that study, hyaluronan had a concen-tration- (0.5, 1 and 2 mg/ml) and molecular weight-dependent (0.5, 1, and 2 kd) inhibition of PGE2 syn-
thesis. The exact molecular weight of each hyaluronanpreparation used in the study reported here is notknown. However, the commercially available hyaluro-nan preparation used in our study is of lower viscositythan other commercially available hyaluronan prepara-tions licensed for intra-articular administration.22
Analysis of the data in the study reported here failed
to confirm the findings of other investigators.17,20 None ofthe 3 hyaluronan preparations, at concentrations up to
Figure 4—Effect of polysulfated glycosaminoglycan (PSGAG) on
1,000 µg/ml, exerted inhibitory actions on LPS-induced
PGE2 synthesis by LPS-stimulated synoviocytes obtained from
PGE2 synthesis. However, at the highest concentration
the metacarpophalangeal joints of clinically normal horses. Eachtreatment group represents mean ± SEM PGE
(2,000 µg/ml), 2 of the 3 preparations (commercially
4 replicate wells. The PSGAG concentrations (in parentheses)
available preparation and S zooepidemicus preparation)
are expressed in µg/ml. See Figure 1 for key.