Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Esm040 367.372
ª The American Genetic Association. 2007. All rights reserved.
For permissions, please email: journals.permissions@oxfordjournals.org.
Fine-Scale Spatial Genetic Structurewithin Continuous and FragmentedPopulations of Trillium camschatcense
HIROKI YAMAGISHI, HIROSHI TOMIMATSU, AND MASASHI OHARA
From the Graduate School of Environmental Earth Science, Hokkaido University, Sapporo 060-0810, Japan (Yamagishi andOhara); and the Department of Biological Sciences, Tokyo Metropolitan University, Hachioji 192-0397, Japan (Tomimatsu).
Address correspondence to M. Ohara at the address above, or e-mail: ohara@ees.hokudai.ac.jp.
Spatial genetic structure (SGS) within populations was ana-
loss of individuals from the family patches are responsible for
lyzed for the ling-lived understory perennial herb Trillium cam-
the juvenile and adult structures, respectively (e.g., Epperson
schatcense using allozyme loci. We used Sp statistics to
and Alvarez-Buylla 1997; Parker et al. 2001; Chung et al. 2003).
compare SGS between 2 life-history stages, juveniles ( J)
Conversely, the opposite trend was also observed in some spe-
and reproductives (R), as well as between 2 populations, con-
cies, and the increasing SGS has been attributed to local selec-
tinuous and fragmented, with different habitat conditions. In
tion during recruitment or historical events (Tonsor et al. 1993;
the continuous population, significant SGS was detected in
Ueno et al. 2000; Kalisz et al. 2001).
both stages but the extent was greatly reduced with the prog-
Moreover, the extent of SGS may vary among popula-
ress of the stage ( J, Sp 5 0.0475; R, Sp 5 0.0053). We in-
tions under different habitat conditions, and such interpop-
ferred that limited seed dispersal and subsequent random
ulation variation can also improve our understanding of the
loss of individuals from the family patches are responsible
relationship between environmental factors and life-history
for the J and R stage structures, respectively. The fragmented
factors. In particular, habitat alteration due to human activ-
population differed in the patterns of SGS; significant struc-
ities affects many demographic and genetic processes includ-
ture was detected in the R stage, but not in the J stage ( J,
ing the breeding system (e.g., Rajimann et al. 1994) and
Sp 5 0.0021; R, Sp 5 0.0165) despite significant positive in-
recruitment (e.g., Tomimatsu and Ohara 2002; Ward and
breeding coefficients ( J, FIS 5 0.251). The observed differen-
Johnson 2005), which are likely reflected in the patterns
ces in the J-stage structures between populations may be
of genetic structuring. Several studies have revealed habi-
explained by habitat fragmentation effects because reduced
tat-associated variations of SGS, for example, between frag-
recruitment in the fragmented population prevents the devel-
mented and continuous populations of Acer saccharum (Young
opment of maternal sibling cohort. Such comparative analysis
and Merriam 1994) and between old-growth and logged for-
between populations and life-history stages can be useful to
ests of Pinus strobus (Epperson and Chung 2001); however,
understand the different underlying causes of SGS.
few studies have examined the differences in SGS of differentlife-history stages both among and within populations (butsee Epperson and Chung 2001; Cruse-Sanders and Hamrick
Spatial genetic structure (SGS) within plant populations is
determined by many processes, including gene flow and local
In this study, we analyzed SGS of 2 life-history stages in 2
selection at different life-history stages (Hedrick 1986; Ennos
populations of the long-lived understory herb, Trillium cam-
1994; Latta et al. 1998; Kalisz et al. 2001). In the case of long-
schatcense Ker Gawler (Trilliaceae or Melanthiaceae). First,
lived perennial plant species, SGS of juveniles primarily reflects
we compared SGS between juveniles and reproductives in
their parental structure and the patterns and distances of
a large, continuous population. Although the species has
both pollen and seed dispersal. This structure can be modified
an outcrossing mating system in our populations (Ohara
in subsequent life-history stages by postdispersal selection or
et al. 1996), the dispersal distance of seeds by ants has been
mortality of individuals. Therefore, analyses of SGS over the
reported to be short, only 0.60 m on the average (Ohara and
life cycle can infer the roles of these demographic and genetic
Higashi 1987). Thus, we expect that the limited seed dispersal
processes in the formation and maintenance of the within-
ability in our species develops significant SGS in juveniles.
population genetic structure (Kalisz et al. 2001; Chung et al.
Juveniles are subsequently subject to density-dependent mor-
2003). For example, some studies have found that significant
tality and many are thus excluded from the family patches
structure in juveniles is greatly reduced in reproductive adults,
(Ohara and Kawano 1986), so that SGS in juveniles is expected
suggesting that limited dispersal and the subsequent random
to decrease or even become undetected in reproductives.
Second, we compared the patterns of SGS in a large popu-
continuous forest (;300 000 m2), whereas the Kiyokawa
lation with those of a small, fragmented population. In the
population (42°45#N, 143°7#E; hereafter ‘‘fragmented’’)
Tokachi plain of eastern Hokkaido, Japan, T. camschatcense
grows in a small, fragmented forest (;7700 m2), fully sur-
has experienced extensive habitat loss because of recent ag-
rounded by agricultural fields. The estimated habitat size
ricultural development since the 1880s and now is often
and the number of reproductive plants are 50 000 m2 and
found in the understory of small and isolated forests. Our
126 000 for the continuous and 7700 m2 and 10 200 for
previous studies (Tomimatsu and Ohara 2002, 2003a) using
the fragmented population, respectively. Our previous study
14 populations with different sizes have demonstrated that
demonstrated that the fragmented population showed a sig-
smaller populations showed lower seed production and seed-
nificant inbreeding coefficient (FIS 5 0.129), possibly due to
ling recruitment and sometimes higher inbreeding coeffi-
biparental inbreeding (Tomimatsu and Ohara 2003a). Con-
cients (i.e., biparental inbreeding). Other things being
sidering the size of habitats and the development history
equal, the reduced recruitment in small populations may pre-
of the region, habitat conditions must be much more altered
vent the development of maternal sibling cohorts and thus
in the fragmented population. Thus, when interpreting the
reduce SGS in juveniles, whereas biparental inbreeding is
data, we assume that environmental conditions that gave
expected to intensify SGS. To examine these expectations,
rise to SGS in the reproductive and juvenile stages are similar
we used spatial autocorrelation analysis with new Sp statistics
in the continuous population, but they are quite different
that has recently been proposed to make a comparative anal-
in the fragmented population owing to the fragmentation
ysis of genetic structure (reviewed by Vekemans and Hardy
2004). Although previous studies used estimators of geneticrelatedness (Moran’s I or kinship coefficients) or the inter-
Leaf Sampling and Allozyme Electrophoresis
cept of spatial correlograms to analyze their variations(e.g., Stehlik and Holderegger 2000; Diniz-Filho and Telles
In this study, we sampled individuals in 2 life-history stages,
2002), these estimators strongly depend on the sampling
reproductives, that is flowering individuals (hereafter ‘‘R’’),
schemes adopted. By contrast, under isolation by distance
and juveniles, that is seedlings and one-leaf individuals (here-
in 2-dimensional space, the Sp statistics primarily depends
after ‘‘J’’). We did not sample 3-leaves nonflowering individ-
on the rate of decrease of pairwise kinship coefficients be-
uals because not enough number of them were found in
tween individuals with the logarithm of the spatial distance
the plots. In each population, leaf materials were collected
in 2 ways. First, to examine the level of genetic diversityand inbreeding coefficient (FIS), leaves were randomly sam-pled from .40 individuals in each life-history stage (R, n 5
41 and J, n 5 94 for the continuous population; R, n 5 52 andJ, n 5 94 for the fragmented population). Second, to analyze
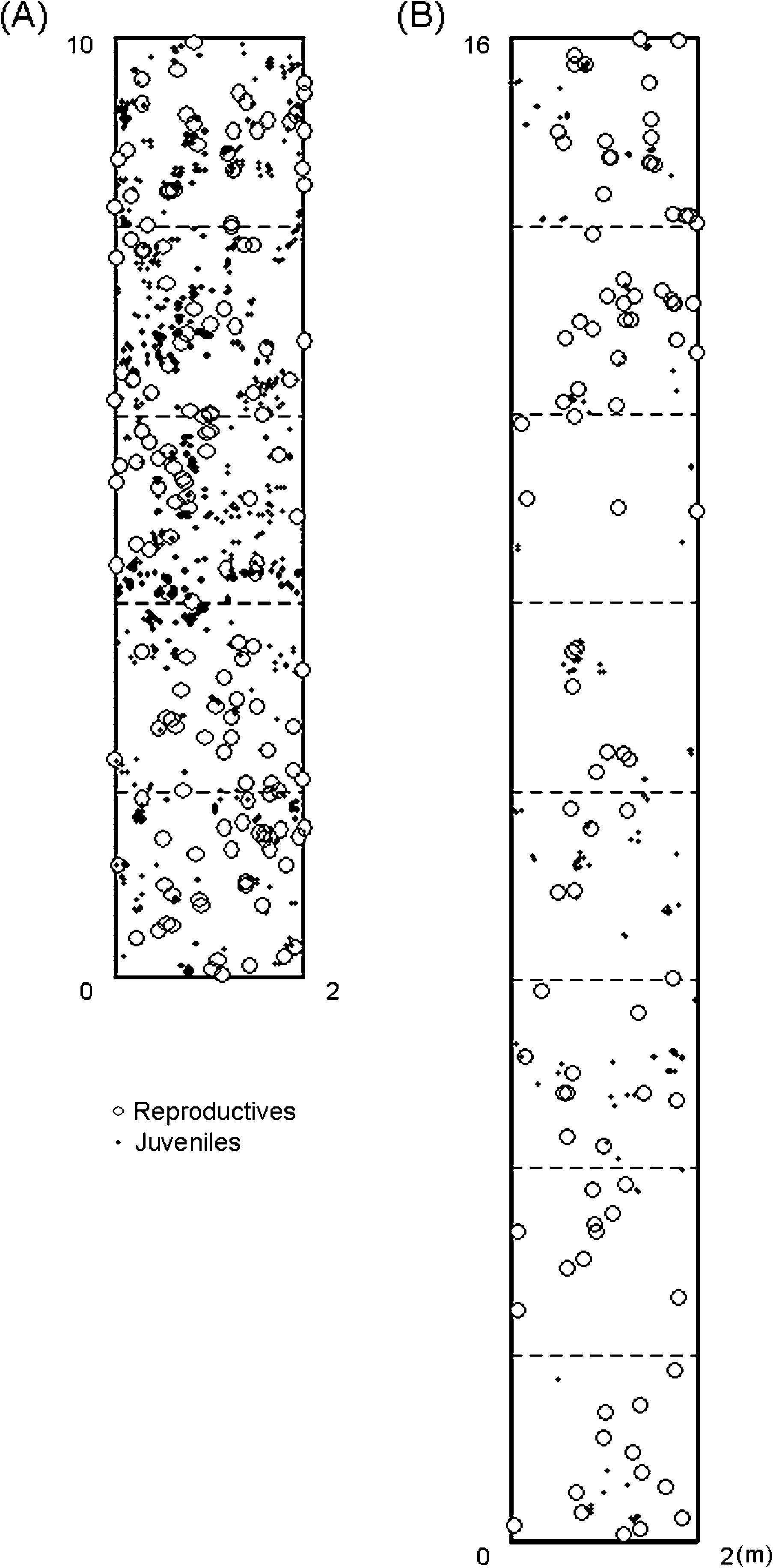
SGS, we established a single plot (2 Â 10 m for the contin-
Trillium camschatcense is a diploid (2n 5 10), nonclonal peren-
uous and 2 Â 16 m for the fragmented; Figure 1) in the center
nial herb that commonly occurs in the understory of broad-
of each population and mapped all individuals of the R and
leaved deciduous forests of Hokkaido, Japan. Vegetative
J stages within the plots. For each stage, we collected leaf
individuals of T. camschatcense comprise 2 morphologically dis-
samples from plants located nearest all intersections of 2 Â
tinct forms: 1-leaf and 3-leaves stages. It takes more than 10
2 cm grids within the quadrants unless there were no plants
years for seedlings to become flowering plants. Reproduc-
within 1 cm of the intersection point. Consequently, we sam-
tives have one or several flowers, and seed production results
pled 151 of R and 754 of J plants in the continuous and 154 of
from obligatory outcrossing by insect pollination in our study
R and 98 of J plants in the fragmented populations. Multi-
area (Ohara et al. 1996). After reaching flowering, the plants
locus genotypes of all sampled individuals were determined
can live for at least 25 years with continuous flowering every
by allozyme electrophoresis as described below. The density
year (Ohara et al. 2006). The flowers are visited by a wide
of R and J plants within the study plots was 7.6 and 71.9 mÀ2
range of insects, primarily beetles (e.g., Nitidulidae and Mel-
in the continuous and 3.1 and 7.8 mÀ2 in the fragmented
andryidae) and flies (e.g., Bibionidae and Scathophagidae;
Tomimatsu and Ohara 2003b), and the fruits contain ;80
Leaf materials were transported on ice to the laboratory
seeds on average (Tomimatsu and Ohara 2002). As each seed
and kept at –80 °C until electrophoresis. Approximately 70
has a soft, juicy elaiosome, the seeds are attractive to ants,
mg of leaf tissue from R plants was homogenized in 0.9 ml
which consequently contribute to seed dispersal (Ohara
of extraction buffer made up of 0.1 M Tris–HCl (pH 8.0),
0.2 g/ml glycerol, 63 mg/ml Tween 80, 8 mM dithiothreitol,
This study was conducted in 2 populations, Hiroo and
0.50% (v/v) b-mercaptoethanol, 0.40% (w/v) b-nicotiamide
Kiyokawa, in the Tokachi plain of eastern Hokkaido, Japan.
adenine dinucleotide, 0.45% (w/v) b-nicotiamide adenine di-
In this study, we selected 2 distinct populations of 14 pop-
nucleotide phosphate, 0.3% (w/v) bovine serum albumin, and
ulations used in our previous studies (Tomimatsu and Ohara
7% (w/v) polyvinylpolypyrrolidone (modified from Shiraishi
2002, 2003a, 2004), one large continuous (Hiroo) and one
1988). As the leaves of J plants were small, we homogenized
small fragmented (Kiyokawa). The Hiroo population
;2 mg of leaf tissue in 0.25 ml of extraction buffer. After the
(42°19#N, 143°20#E; hereafter ‘‘continuous’’) grows in a large,
homogenates were centrifuged (15 000 rpm for 15 min at
Standard measures of genetic diversity, including the percent-age of polymorphic loci (P), the mean number of alleles perlocus (A) corrected for differences in sample size (i.e., rare-fied; Leberg 2002), the observed heterozygosity (Ho), and theexpected heterozygosity (He) were calculated for each life-history stage in each population. Inbreeding coefficient(FIS) was calculated as estimates of Weir and Cockerham(1984) using the computer program FSTAT version2.9.3.2 (Goudet 2002), and the levels of significance wereobtained by randomization-based procedures (for details,see Goudet 2002). To investigate fine-scale SGS, we con-ducted spatial autocorrelation analyses with kinship coeffi-cients (Loiselle et al. 1995) using SPAGeDi version 1.2(Hardy and Vekemans 2002). Mean multilocus kinship coef-ficients (F( j )) were computed for the following distance clas-ses: 0.2, 0.3, 0.4, 0.5, 0.75, 1, 2, 4, and 8 (upper-bounddistance in meters) and were plotted against the logarithmof the geographic distance ( j ). Standard errors for the kinshipcoefficients were estimated using a jackknife procedure overthe loci. The linear regression slope (b) can be a good esti-mator of the extent of SGS (Vekemans and Hardy 2004). We tested the significance of b against the null hypothesisH0: b 5 0 (i.e., the overall absence of SGS) by comparingthe observed values with those obtained after 1000 randompermutations of individuals among positions. We also quan-tified SGS by Sp statistics, calculated as Àb/(1 À F(1)) whereF(1) is the mean kinship coefficient in the first distance class(see Vekemans and Hardy 2004 for details). The Sp statisticscould be considered robust in our sampling scheme becauseF( j ) tended to decrease linearly with ln( j ).
Genetic Diversity and Levels of Inbreeding
Seven of the 8 loci resolved were polymorphic in at leastone of the 2 populations examined: 6 (AAT [2 loci],ADH, SODH, LAP, GDH) and 4 (AAT [2 loci], ADH,LAP) loci were polymorphic in the continuous and frag-mented populations, respectively; MDH was monomorphic.
Spatial distribution of Trillium camschatcense
In both stages, the continuous population had higher genetic
individuals within the study plots. Individuals of different stage
diversity (P, A, Ho, and He) than the fragmented population
classes are represented by different symbols in the maps. Maps
(Table 1). For example, when the differences in sample sizes
are shown for (A) continuous and (B) fragmented populations.
were adjusted, the mean number of alleles per locus (A) was1.875 for R and 2.227 for J stages in the continuous popu-lation and 1.620 for R and 1.484 for J stages in the fragmented
4 °C), 10 ll of the resulting supernatant was used for vertical
population. In the continuous population, inbreeding co-
polyacrylamide gel electrophoresis. We tested 14 enzymes
efficients were nearly equal to zero (R, FIS 5 0.072; J,
first, and then examined 6 enzyme systems that consistently
FIS 5 0.079). In contrast, the fragmented population showed
showed clear and interpretable banding patterns: aspartate
high inbreeding coefficients in both stages (R, FIS 5 0.189;
aminotransferase (AAT, 2 loci [EC 2.6.1.1]), leucine amino-
J, FIS 5 0.251). Randomization tests detected a significant
peptidase (LAP, 1 locus [EC 3.4.11.1]), glutamate dehydroge-
deficit of heterozygosity relative to Hardy–Weinberg expect-
nase (GDH, 1 locus [EC 1.4.1.2]), malate dehydrogenase
(MDH, 1 locus [EC 1.1.1.37]), sorbitol dehydrogenase
(SODH, 1 locus [EC 1.1.1.14]), and alcohol dehydrogenase(ADH, 2 loci [EC 1.1.1.1]). On the basis of the banding pat-
The slopes (b) of linear regressions between the kinship coef-
terns, we assumed Mendelian inheritance of all loci.
ficient and the logarithm of the geographic distance were
Genetic diversity and SGS of continuous and fragmented populations of Trillium camschatcense
Genetic diversity and inbreeding coefficient
P, percent polymorphic loci; A, average number of alleles per locus corrected for differences in sample size; Ho, observed heterozygosity; He, expectedheterozygosity (gene diversity); FIS, estimates of Weir and Cockerham (1984) of inbreeding coefficient; Sp, estimates of Vekemans and Hardy (2004) ofSGS; b, slope of the regression of kinship coefficients on the logarithm of geographic distance (*P , 0.05; **P , 0.01); F(1), the value of F(r) on the firstdistance class.
found to be significantly negative (P , 0.05) in all but the J
tion exhibited significant SGS at the R stage (Sp 5 0.0165), but
stage in the fragmented population (Table 1; Figure 2). In the
not in the J stage (Sp 5 0.0021). When the strength of SGS of
continuous population, the Sp statistics showed that the J stage
each life-history stage was compared between populations,
was much more genetically structured than the R stage (J, Sp 5
the R stage in the fragmented population was slightly more
0.0475; R, Sp 5 0.0053). In contrast, the fragmented popula-
structured than that in the continuous population (continuous,
Correlograms (solid lines) resulting from spatial autocorrelation analyses based on allozyme loci for 2 study
populations of Trillium camschatcense. Least-squares linear regressions are indicated by dotted lines. Vertical bars show standarderrors. Distance classes are 0.2, 0.3, 0.4, 0.5, 0.75, 1, 2, 4, and 8 m (upper-bound distance).
Sp 5 0.0053; fragmented, Sp 5 0.0165). In contrast, the J stage
This suggests that only a few maternal siblings exist among
exhibited about 20-fold stronger structure in the continuous
juveniles, and thus the low recruitment could explain the ab-
population than in the fragmented population (continuous,
sence of genetic structure. Interestingly, a related species, T.
Sp 5 0.0475; fragmented, Sp 5 0.0021).
grandiflorum, also did not show significant SGS at the juvenilestage (Kalisz et al. 2001). The number of seeds produced perreproductive plant is small in T. grandiflorum (;16 seeds; Kalisz
et al. 1999), so that the resulting low recruitment, in conjunc-
This study indicates how fine-scale SGS of T. camschatcense
tion with a moderate level of seed dispersal, would have a ho-
differs across 2 life-history stages and 2 populations under
mogenizing effect on the genetic structure (Kalisz et al. 2001).
contrasting habitat conditions. In addition, the fragmented
Although the difference between Sp statistics was not tested in
population exhibited lower allelic diversity and higher
our study, the reproductive stage SGS in the fragmented pop-
inbreeding coefficients than the continuous population
ulation was slightly stronger with lower adult density than that
(Table 1). These results are consistent with our previous
in the continuous population (Table 1; Figure 2). This result is
study (Tomimatsu and Ohara 2003a). The fragmented pop-
consistent with theoretical and empirical evidence that density
ulation would have experienced a stochastic loss of alleles at
is a major determinant of SGS because it affects the strength
the time of fragmentation and biparental inbreeding due to
of local genetic drift and the level of gene flow distance (Hey-
wood 1991; Vekemans and Hardy 2004). We are not sure,
In the continuous population, as expected, we found sig-
however, whether this difference in SGS is significant because
nificant SGS in juveniles (Table 1; Figure 2A). This structure
F(1) in these populations are similar (Table 1) and F( j) in the
could develop as a result of limited pollen and seed dispersal
short distance classes had large variance in the fragmented
(Wright 1946; Slatkin 1985). Although the distance of pollen
dispersal would not be far, the value of inbreeding coefficient
Some limitations and caveats should be considered before
(FIS) indicates that mating within this population was essen-
we draw any conclusions from this study. First, we examined
tially random (Table 1). Therefore, the results suggest that re-
only a single plot within each population. Although we care-
stricted seed dispersal is mainly responsible for SGS in
fully established the plots to reflect typical environmental
juveniles. In fact, seeds that fell on the ground were rarely
conditions and demographic structures of populations, 2
transported by ants even after 72 h and many remained around
or more replicates are needed to draw more corroborative
the maternal plants (dispersal frequency 11.7%; Yamagishi H,
evidence. Second, the accuracy of Sp statistics critically
unpublished data). By contrast, the reproductive stage showed
depends on F( j ) in the first several distance classes, particu-
a weak structure relative to the juvenile stage (Table 1; Figure
larly F(1) (Vekemans and Hardy 2004). For example, F( j ) in
2A). The reduction pattern of SGS with the progress of the
the first 4 distance classes had large standard errors in the con-
stage could occur when juveniles experience genetically ran-
tinuous population (Figure 2A) and this could partly be
dom mortality that occurs in a density-dependent manner.
caused by how we defined distance classes. We recalculated
Several previous studies also demonstrated the same pattern,
F(1) and Sp statistics under different sets of distance classes,
which was attributed to random mortality during recruitment
but these calculations gave essentially the same interpretation
(Hamrick et al. 1993; Epperson and Alvarez-Buylla 1997;
of our data (results not shown). Finally, in the fragmented
Chung et al. 2003). It should be noted, however, that the
population, we do not infer the causes of creating the strong
opposite trend was observed in a related species of the same
pattern of structure observed in the reproductives from the
genus, Trillium grandiflorum (Kalisz et al. 2001); significant SGS
pattern in the juveniles. Considering the long life-span of T.
was detected in reproductive but not in juvenile stages, and it
camschatcense (.50 years) and the population history, many
was suggested that the increase in SGS between the stages
reproductive individuals may still have survived fragmenta-
could arise from local selection resulting from microenviron-
tion. Therefore, the existent structure in this reproductive
mental conditions or from specific historical events.
population was likely to develop under habitat conditions be-
The fragmented population differs in the extent and pat-
fore or immediately after habitat fragmentation, whereas the
tern of SGS from the continuous population; namely, juve-
structure in the juveniles was established recently, long after
niles did not show significant SGS despite relatively high
levels of inbreeding coefficients (Table 1; Figure 2B). Given
In summary, under a high level of recruitment, SGS
our previous knowledge on the effects of habitat fragmenta-
appears at the juvenile stage probably because of limited seed
tion, juvenile stage SGS could either increase due to biparental
dispersal around maternal plants, and subsequent random
inbreeding or decrease due to reduced recruitment. Our results
loss of individuals from family patches is a plausible explana-
suggest that the low recruitment may be responsible for the
tion for the loss of SGS from reproductive to juvenile stages.
absence of SGS. In this population, seedling recruitment is
However, under low recruitment due to habitat fragmenta-
much more restricted than in the continuous population prob-
tion, juveniles do not show a significant genetic structure pos-
ably because of reduced seed production, microclimatic edge
sibly because only few maternal siblings exist. Although
effects, and biparental inbreeding (Tomimatsu and Ohara
replicates of plots or populations may be needed to confirm
2002, 2004). Actually, the frequency of juveniles was only
our conclusions, such comparative analysis between popula-
2.5 times that of reproductives in the fragmented population
tions as well as between life-history stages can be useful to
(cf., 9.5 times that of the continuous population; Figure 1).
understand the different potential causes of the SGS.
Ohara M, Higashi S. 1987. Interference by ground beetles with the dispersalby ants of seeds of Trillium species (Liliaceae). J Ecol. 75:1091–1098.
The Akiyama Memorial Foundation; the Foundation for
Ohara M, Kawano S. 1986. Life history studies on the genus Trillium (Lil-
Earth Environment; the Ministry of Education, Culture,
iaceae) IV. Stage class structures and spatial distribution of four Japanese
Sports, Science; and Technology for the 21st Century Center
species. Plant Species Biol. 1:147–161.
of Excellence Program (E-01) and the Japan Society for the
Ohara M, Takeda H, Ohno Y, Shimamoto Y. 1996. Variations in the breed-
Promotion of Science for Scientific Research (Nos. 16370007
ing systems and the population genetic structure of Trillium kamtschaticum (Lil-
Ohara M, Tomimatsu H, Takada T, Kawano S. 2006. Importance of life his-tory studies for conservation of fragmented populations: a case study of the
understory herb, Trillium camschatcense. Plant Species Biol. 21:1–12.
We are grateful to K. Nemoto for assistance in the field and Hiroo town
Parker KC, Hamrick JL, Parker AJ, Nason JD. 2001. Fine-scale genetic struc-
office for offering the use of their facility. We also thank M. T. Kimura
ture in Pinus clausa (Pinaceae) populations: effects of disturbance history. He-
Rajimann LEL, van Leeuwen NC, Kersten R, Oostermeijer JGB, den NijsHCM, Menken SBJ. 1994. Genetic variation and outcrossing rate in re-
lation to population size in Gentiana pneumonanthe L. Conserv Biol. 8:
Chung MY, Epperson BK, Chung MG. 2003. Genetic structure of age clas-
ses in Camellia japonica (Theaceae). Evolution. 57:62–73.
Shiraishi S. 1988. Inheritance of isozyme variations in Japanese black pine,
Cruse-Sanders JM, Hamrick JL. 2004. Spatial and genetic structure within
Pinus thunbergii Parl. Silvae Genet. 37:93–100.
populations of wild American ginseng (Panax quinquefolius L., Araliaceae).
Slatkin M. 1985. Gene flow in natural populations. Annu Rev Ecol Syst.
Diniz-Filho JA, Telles MPC. 2002. Spatial autocorrelation analysis and the
Stehlik I, Holderegger R. 2000. Spatial genetic structure and clonal diversity
identification of operational units for conservation in continuous popula-
of Anemone nemorosa in late successional deciduous woodlands of Central
Ennos RA. 1994. Estimating the relative rates of pollen and seed migration
Tomimatsu H, Ohara M. 2002. Effects of forest fragmentation on seed
among plant populations. Heredity. 72:250–259.
production of the understory herb Trillium camschatcense. Conserv Biol.
Epperson BK, Alvarez-Buylla ER. 1997. Limited seed dispersal and genetic
structure in life stages of Cecropia obtufifolia. Evolution. 51:275–282.
Tomimatsu H, Ohara M. 2003a. Genetic diversity and local population struc-
Epperson BK, Chung MG. 2001. Spatial genetic structure of allozyme poly-
ture of fragmented populations of Trillium camschatcense (Trilliaceae). Biol
morphisms within populations of Pinus strobus (Pinaceae). Am J Bot.
Tomimatsu H, Ohara M. 2003b. Floral visitors of Trillium camschatcense (Tril-
Goudet J. 2002. FSTAT, a program to estimate and test gene diversities and
liaceae) in fragmented forests. Plant Species Biol. 18:123–127.
fixation indices, version 2.9.3.2 [Internet]. [cited 2005 Oct 1]. Available from:
Tomimatsu H, Ohara M. 2004. Edge effects on recruitment of Trillium cam-
schatcense in small forest fragments. Biol Conserv. 117:509–519.
Hamrick JL, Murawski D, Nason JD. 1993. The influence of seed dispersal
Tonsor SJ, Kalisz S, Fisher J. 1993. A life-history based study of popula-
mechanisms on the genetic structure of tropical tree populations. Vegetatio.
tion structure: seed bank to adults in Plantago lanceolata. Evolution. 47:
Hardy OJ, Vekemans X. 2002. SPAGeDi: a versatile computer program to
Ueno S, Tomaru N, Yoshimaru H, Manabe T, Yamamoto S. 2000. Genetic
analyse spatial genetic structure at the individual or population levels. Mol
structure of Camellia japonica L. in an old-growth evergreen forest, Tsushima,
Hedrick PW. 1986. Genetic polymorphism in heterogeneous environments:
Vekemans X, Hardy OJ. 2004. New insights from fine-scale spatial genetic
a decade later. Annu Rev Ecol Syst. 17:235–255.
structure analyses in plant populations. Mol Ecol. 13:921–935.
Heywood JS. 1991. Spatial analysis of genetic variation in plant populations.
Ward M, Johnson SD. 2005. Pollen limitation and demographic structure in
small fragmented populations of Brunsivigia radulosa (Amaryllidaceae). Oikos.
Kalisz S, Hamzawa FM, Tonsor SJ, Thiede DA, Voigt S. 1999. Ant-mediated
seed dispersal alters pattern of relatedness in a population of Trillium gran-
Weir BS, Cockerham CC. 1984. Estimating F-statistics for analysis of pop-
ulation structure. Evolution. 43:413–418.
Kalisz S, Nason JD, Hanzawa FM, Tonsor SJ. 2001. Spatial population ge-
Wright S. 1946. Isolation by distance under diverse systems of mating. Ge-
netic structure in Trillium grandiflorum: the roles of dispersal, mating, history,
and selection. Evolution. 55:1560–1568.
Young AG, Merriam HG. 1994. Effects of forest fragmentation on the spa-
Latta RG, Linhart YB, Fleck D, Elliot M. 1998. Direct and indirect estimates
tial genetic structure of Acer saccharum Marsh. (sugar maple) populations. He-
of seed versus pollen movement within a population of ponderosa pine. Evo-
Leberg PL. 2002. Estimating allelic richness: effects of sample size and bot-
Loiselle BA, Sork VL, Nason JD, Graham C. 1995. Spatial genetic structureof a tropical understory shrub. Am J Bot. 82:1420–1425.
Transmission Symptoms Treatment Reduce Your Risk prevents the most common strains of HPV. can’t see a sore during penis, vagina, anus, Blood, semen, vaginal Possibly no symptoms. Blood, semen, vaginal Depleted immune system. No cure. infections in 2006. enters the body of an If you want to be protected against STIs, practice safer
 Standard measures of genetic diversity, including the percent-age of polymorphic loci (P), the mean number of alleles perlocus (A) corrected for differences in sample size (i.e., rare-fied; Leberg 2002), the observed heterozygosity (Ho), and theexpected heterozygosity (He) were calculated for each life-history stage in each population. Inbreeding coefficient(FIS) was calculated as estimates of Weir and Cockerham(1984) using the computer program FSTAT version2.9.3.2 (Goudet 2002), and the levels of significance wereobtained by randomization-based procedures (for details,see Goudet 2002). To investigate fine-scale SGS, we con-ducted spatial autocorrelation analyses with kinship coeffi-cients (Loiselle et al. 1995) using SPAGeDi version 1.2(Hardy and Vekemans 2002). Mean multilocus kinship coef-ficients (F( j )) were computed for the following distance clas-ses: 0.2, 0.3, 0.4, 0.5, 0.75, 1, 2, 4, and 8 (upper-bounddistance in meters) and were plotted against the logarithmof the geographic distance ( j ). Standard errors for the kinshipcoefficients were estimated using a jackknife procedure overthe loci. The linear regression slope (b) can be a good esti-mator of the extent of SGS (Vekemans and Hardy 2004).
Standard measures of genetic diversity, including the percent-age of polymorphic loci (P), the mean number of alleles perlocus (A) corrected for differences in sample size (i.e., rare-fied; Leberg 2002), the observed heterozygosity (Ho), and theexpected heterozygosity (He) were calculated for each life-history stage in each population. Inbreeding coefficient(FIS) was calculated as estimates of Weir and Cockerham(1984) using the computer program FSTAT version2.9.3.2 (Goudet 2002), and the levels of significance wereobtained by randomization-based procedures (for details,see Goudet 2002). To investigate fine-scale SGS, we con-ducted spatial autocorrelation analyses with kinship coeffi-cients (Loiselle et al. 1995) using SPAGeDi version 1.2(Hardy and Vekemans 2002). Mean multilocus kinship coef-ficients (F( j )) were computed for the following distance clas-ses: 0.2, 0.3, 0.4, 0.5, 0.75, 1, 2, 4, and 8 (upper-bounddistance in meters) and were plotted against the logarithmof the geographic distance ( j ). Standard errors for the kinshipcoefficients were estimated using a jackknife procedure overthe loci. The linear regression slope (b) can be a good esti-mator of the extent of SGS (Vekemans and Hardy 2004).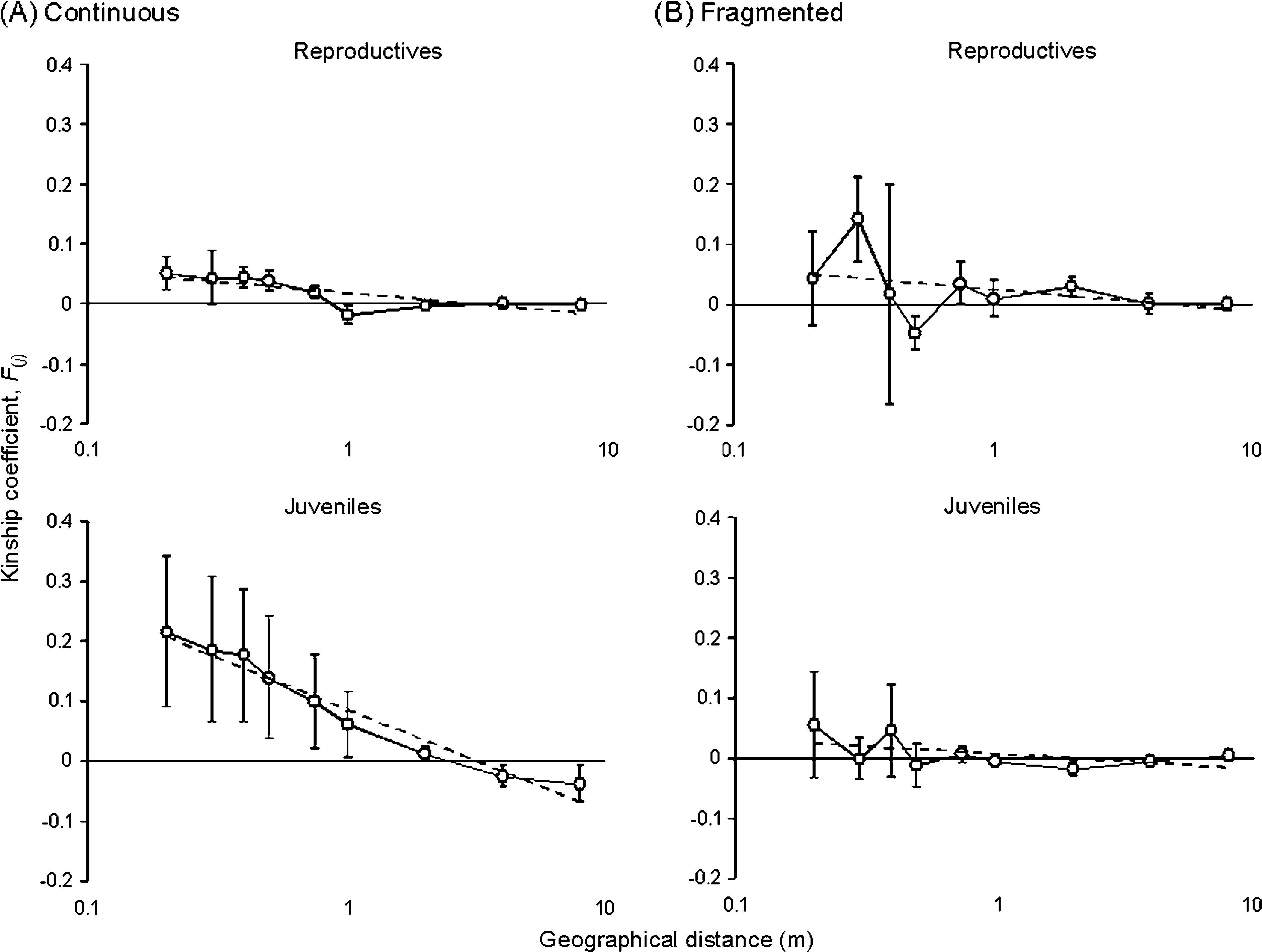 Genetic diversity and SGS of continuous and fragmented populations of Trillium camschatcense
Genetic diversity and inbreeding coefficient
P, percent polymorphic loci; A, average number of alleles per locus corrected for differences in sample size; Ho, observed heterozygosity; He, expectedheterozygosity (gene diversity); FIS, estimates of Weir and Cockerham (1984) of inbreeding coefficient; Sp, estimates of Vekemans and Hardy (2004) ofSGS; b, slope of the regression of kinship coefficients on the logarithm of geographic distance (*P , 0.05; **P , 0.01); F(1), the value of F(r) on the firstdistance class.
Genetic diversity and SGS of continuous and fragmented populations of Trillium camschatcense
Genetic diversity and inbreeding coefficient
P, percent polymorphic loci; A, average number of alleles per locus corrected for differences in sample size; Ho, observed heterozygosity; He, expectedheterozygosity (gene diversity); FIS, estimates of Weir and Cockerham (1984) of inbreeding coefficient; Sp, estimates of Vekemans and Hardy (2004) ofSGS; b, slope of the regression of kinship coefficients on the logarithm of geographic distance (*P , 0.05; **P , 0.01); F(1), the value of F(r) on the firstdistance class.