Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Wang.salk.edu
[12] The side product related to 6 was also formed, but the optical purity
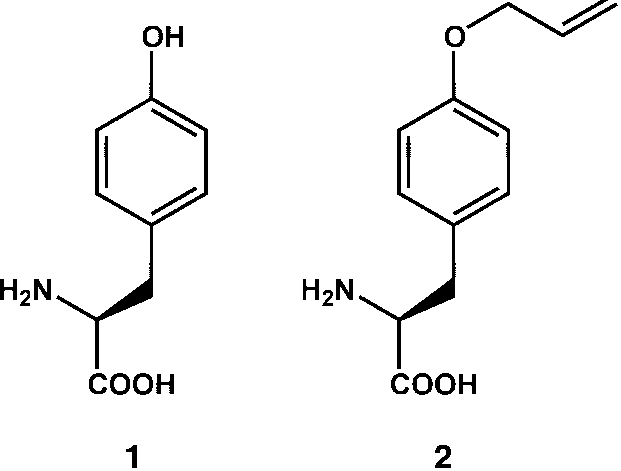
of O-allyl-l-tyrosine (2) into proteins in E. coli. The alkene
functional group of this unnatural amino acid should provide
[13] M. Shimazaki, H. Haram, K. Suzuki, G.-i. Tsuchihashi, Tetrahedron
new chemical methods for the selective modification of
Lett. 1987, 28, 5891; K. Suzuki, E. Katayama, G.-i. Tsuchihashi,
[14] Rearrangements of optically active vinyl epoxides mediated by Lewis
Previously we generated an orthogonal tRNA ± synthetase
acids give quaternary asymmetric carbon centers with inversion of the
pair, mutRNATyr ± TyrRS, in E. coli by modifying the
configuration: a) M. E. Jung, D. C. D×Amino, J. Am. Chem. Soc. 1995,
tRNATyr ± TyrRS pair of Methanococcus jannaschii.[4, 5] This
117, 7379; b) for rearrangements that involve a sterically restricted
mutRNATyr is not aminoacylated by endogenous synthetases
cation center, see: S. A. Monti, S.-C. Chen, Y.-L. Yang, S.-S. Yuan,
O. P. Bourgeois, J. Org. Chem. 1978, 43, 4062.
in E. coli, and functions well in translation. The TyrRS does
[15] The enantiomeric purity of 15 was determined by HPLC (Chiralcel-
not aminoacylate E. coli tRNAs,[6] but aminoacylates the
with tyrosine (1); the acylated mutRNATyr
[16] C. H. Senanayake, R. D. Larsen, T. J. Bill, J. Liu, E. G. Corley, P. J.
inserts tyrosine in response to the amber nonsense codon. To
change the substrate specificity of the TyrRS so that itaminoacylates the mutRNATyr
common amino acids, a mutant TyrRS library was generated
The Selective Incorporation of Alkenes intoProteins in Escherichia coli**
Zhiwen Zhang,[] Lei Wang,[] Ansgar Brock, andPeter G. Schultz*
The addition of amino acids with novel functional groups to
the genetic code of Escherichia coli should greatly enhanceour ability to study protein structure and function, as well asgenerate proteins with novel properties. We recently showedthat the unnatural amino acids O-methyl-l-tyrosine and l-3-
and selected. Based on an analysis of the crystal structure of
(2-naphthyl)alanine can be site-specifically incorporated into
the homologous TyrRS from Bacillus stearothermophilus,[7]
proteins in Escherichia coli with high efficiency and fidel-
five residues (Tyr 32, Glu 107, Asp 158, Ile 159, and Leu 162) in
ity.[1, 2] This result requires the addition of an orthogonal
the active site of M. jannaschii TyrRS that are within 6.5 ä of
tRNA ± codon pair and aminoacyl-tRNA synthetase to the
the para position of the aryl ring of tyrosine were randomly
translational machinery of the cell. The new synthetase (and
mutated.[1, 8] This mutant library was first subjected to a
only this synthetase) aminoacylates the orthogonal tRNA
positive selection based on the suppression of an amber codon
(and only this tRNA) with the unnatural amino acid only,
introduced at a nonessential position (Asp 112) in the
which is inserted into proteins in response to the amber
chloramphenicol acetyl transferase (CAT) gene. Cells trans-
codon, TAG.[3] We report here the site-specific incorporation
formed with the mutant TyrRS libraries, the mutRNATyr
gene, and the amber mutant CAT gene were grown in minimal
media containing 1 mm 2 and 70 mg mLÀ1 chloramphenicol.
The survivors must encode a mutant TyrRS that amino-
the Skaggs Institute for Chemical Biology
acylates the mutRNATyr with either 2 or endogenous amino
acids. To remove mutant synthetases with specificities for
10550 North Torrey Pines Road, La Jolla, CA 92037(USA)
endogenous amino acids, a negative selection was applied.
Fax: ( 1) 858-784-9440E-mail: schultz@scripps.edu
Three amber codons were introduced at nonessential posi-
tions (Gln 2, Asp 44, Gly 65) in the toxic barnase gene.[9] Cells
expressing the mutant synthetase from the positive selection,
the mutRNATyr gene, and the amber mutant barnase gene
10550 North Torrey Pines Road, La Jolla, CA 92037(USA)
were grown in Luria ± Bertani (LB) media in the absence of 2.
Cells encoding synthetases with specificities for any endoge-
Department of ChemistryUniversity of California at Berkeley
nous amino acids will produce barnase and die. Only those
encoding a mutant synthetase with specificity for 2 can
Genomics Institute of the Novartis Research Foundation
After three rounds of positive selection alternating with two
10675 John Jay Hopkins Drive, San Diego, CA 92121 (USA)
rounds of negative selection, a clone was evolved whose
[] These authors contributed equally to this work.
survival in chloramphenicol was dependent on the presence of
[**] Support for this work was provided by the Department of Energy,
2 when the selected mutant TyrRS gene (AL-TyrRS) was
Basic Energy Sciences Program, and the National Institutes of Health.
coexpressed with the Asp 112amber CAT gene and the
We thank Dr. Steve Santoro for plasmid pREP(2) and Sheng Ding forhelpful discussions. This is TSRI manuscript no. 14950-CH.
mutRNATyr gene. Cells can survive in 120 mg mLÀ1 chloram-
¹ WILEY-VCH Verlag GmbH, 69451 Weinheim, Germany, 2002
phenicol in the presence of 2, and up to 10 mg mLÀ1
chloramphenicol in the absence of 2. For comparison, E. coli
mutRNATyr ± AL-TyrRS was analyzed by electrospray ion-
cells expressing the Asp112amber CAT gene and the
ization Fourier transform ion cyclotron resonance mass
gene survive in 4 mg mLÀ1 chloramphenicol.[5]
spectrometry (FT-ICR MS). The experimental monoisotopic
This result suggests that the AL-TyrRS has higher activity for
mass of the intact protein was 7963.891 Da, which is within
2 than for natural amino acids. The evolved AL-TyrRS has the
1 ppm of the theoretical mass of 7963.889 Da. Another major
following mutations: Glu 107 3Ala 107, Asp 158 3Cys 158,
signal corresponds to the protein without the first methionine
and Ile 159 3Ala 159. The residues Tyr 32 and Leu 162 remain
unchanged. The mutations of Glu 107 3Ala 107and Ile 159
The signal-to-noise ratio of more than 1500 observed in the
3Ala 159 may enlarge the active site of the mutant synthetase
intact protein mass spectrum suggests a fidelity for the
to accommodate the allyl group. The exact roles of these
incorporation of 2 of better than 99.8 %. This result clearly
mutations will be revealed by solving the crystal structure of
demonstrates the site-specific incorporation of 2 in response
to the amber codon by the mutRNATyr ± AL-TyrRS, and that
To confirm that the observed phenotype is caused by the
other endogenous E. coli synthetases do not utilize 2 as a
site-specific incorporation of 2 by the mutRNATyr ± AL-
TyrRS pair, a mutant Z domain[10, 11] protein was produced
In summary, a useful nonnatural amino acid, O-allyl-l-
and characterized. An amber codon was introduced at the
tyrosine, has been site-specifically incorporated into proteins
seventh position in the gene encoding the Z domain. A His6
in vivo. The allyl group is versatile in organic transformations,
tag was added to the C terminus of the Z domain to facilitate
including metathesis,[13] Diels-Alder and 1,3-dipolar cyclo-
protein purification by Ni2-affinity chromatography. As a
addition reactions.[14] Olefin metathesis has been used suc-
positive control, the wild type M. jannaschii TyrRS was
cessfully to cross-link amino acid derivatives[15] and cyclize
coexpressed with the mutRNATyr , which resulted in the
peptides.[16, 17] Recent progress in developing water-soluble
suppression of the amber codon with tyrosine and production
ruthenium catalysts[18] should facilitate the application of this
of full-length Z-domain protein (Figure 1). When AL-TyrRS
reaction to the modification of proteins containing alkenegroups.[19, 20] Moreover, the allyl side chain of this nonnaturalamino acid may confer new physical properties on proteins aswell.
All chemicals were purchased from Aldrich. NMR spectroscopic data wasrecorded using a Bruker AMX 400. Mass spectra of small molecules wereobtained from Scripps Center for Mass Spectrometry.
Synthesis of 2: O-allyl-l-tyrosine (2) was synthesized according to thepublished procedures[21] with minor modifications. N-(tert-butoxycarbon-yl)-l-tyrosine (2.95 g, 10 mmole) was dissolved in 80 mL of N,N-dimethyl-formamide (DMF). The solution was cooled to 5 8C and NaH (0.63 g,26 mmole) was added. The reaction mixture was allowed to warm up to10 8C and stirred for an additional 2 h. Allyl bromide (1.33 g, 11 mmole)was then added with stirring and the reaction mixture was warmed to roomtemperature and stirred for an additional 4 h. Water was added and theaqueous layer was extracted with ethyl acetate and CH2Cl2. The organiclayer was dried over anhydrous MgSO
Figure 1. A) Amino acid sequence of the Z domain. X indicates the
to afford a white solid, which was then refluxed in 4 m HCl in 1,4-dioxane
position for incorporation of 2 encoded by the amber nonsense codon;
for 4 h. All solvent was evaporated to give the desired product as a white
B) SDS-PAGE analysis of the accumulation of Z domain under different
conditions. The far left lane is a molecular-weight marker. Expression
5.1 Hz, 1 H), 4.53 (d, J 4.6 Hz, 2 H), 5.37(q, J 17.4, 11.3, 10.6 Hz, 1 H),
conditions are noted at the top of each lane. Proteins were purified by Ni2-
5.99 (m, 1 H), 6.91 (d, J 8.4 Hz, 2 H), 7.12 ppm (d, J 8.4 Hz, 2 H);[21]
affinity chromatography and the gel was silver-stained.[12] AL-TyrRS
O-allyl-l-tyrosine-specific mutant synthetase; wt wild type.
3OD): d 40.0, 69.5, 73.1, 115.8, 117.4, 130.5, 131.5, 135.6,
158.4, 177.5 ppm; m/z (ESI, MH): 222.19.
Selection for AL-TyrRS: The positive selection based on the suppression ofan amber codon in chloramphenicol acetyl transferase was carried out as
was expressed together with the mutRNATyr in the presence
described.[1] For the negative selection, plasmid pLWJ17B3 was used to
of 2, full-length Z domain was also obtained. In the absence of
under the control of the lpp promoter and
either the AL-TyrRS, the mutRNATyr , or 2, no full-length
rrnC terminator, and the barnase gene with three amber codons at Gln 2,
Asp 44, and Gly 65 under arabinose induction. After positive selection,
Z domain was observed. These results show that full-length
pBK plasmids encoding mutant TyrRS were isolated and transformed into
mutant protein is produced only in the presence of the AL-
E. coli DH10B competent cells harboring pLWJ17B3. Cells were grown in
TyrRS, the mutRNATyr , and 2. The yield of full-length
LB media containing 0.2 % arabinose, 50 mg mLÀ1 kanamycin, and
mutant Z domain containing 2 is 5.6 mg LÀ1 in minimal media.
35 mg mLÀ1 chloramphenicol. After 8 h, cells were pelleted, and pBKplasmids were purified for further rounds of selection. After sequential
For comparison, the yield of Z-domain is 9.2 mg LÀ1 when the
positive, negative, positive, negative, then positive selection, the candidate
and wild-type M. jannaschii TyrRS are coex-
pBK-ALRS encoding AL-TyrRS was identified and characterized using an
in vivo chloramphenicol acetyl transferase assay.[5]
¹ WILEY-VCH Verlag GmbH, 69451 Weinheim, Germany, 2002
Protein expression, purification, and characterization: Plasmid pLEIZ was
Synthesis andCharacterization of the Neutral
used to express the Z-domain gene with an amber codon at the seventh
position under the control of a bacteriophage T5 promoter and t
nator, and the mutRNATyr gene under the control of the lpp promoter and
Na2Ar'GaGaAr' (Ar' 2,6-Dipp2C6H3, Dipp
rrnC terminator. The AL-TyrRS gene was encoded in plasmid pBK-ALRS
under the control of the constitutive E. coli GlnRS promoter andterminator. E. coli DH10B cells cotransformed with pLEIZ and pBK-
Ned J. Hardman, Robert J. Wright, Andrew D. Phillips,
ALRS were grown in minimal media containing 1 % glycerol and 0.3 mm
leucine (GMML media) with 25 mg mLÀ1 kanamycin, 34 mg mLÀ1 ofchloramphenicol, and 0.5 mm 2. When cells reach an optical density
The publication of the remarkable gallium compound
(OD600) value of 0.5, isopropyl-b-d-thiogalactopyranoside (IPTG; 1 mm)was added to induce protein expression. After 4 h, cells were pelleted and
the protein was purified by Ni2-affinity chromatography according to the
iPr3C6H2) in 1997resulted in much controversy owing to the
manufacturer×s protocol under denaturing conditions (Quiagen, Valencia,
claim that it contained a GaÀGa triple bond.[1] Initially, the
CA). Proteins were then desalted by using a PD-10 column (Amersham
triple bonding in this molecule was justified on the basis of a
Pharmacia, Piscataway, NJ) and eluted in water. The yield of protein was
short GaÀGa distance (2.319(3) ä) and the correspondence of
measured by Bradford assay (BCA kit, Biorad, Hercules, CA). Aliquots ofprotein were used for SDS-PAGE and mass spectroscopic analysis.
the putative [Ar*GaGaAr*]2À ion to the neutral germaniumspecies Ar*GeGeAr*–a germanium ± alkyne analogue. Al-
though the existence of GaÀGa triple bonding has receivedsupport from some calculations,[2±6] others have questionedthis view on the basis of 1) the trans-bent structure of the Cipso-
[1] L. Wang, A. Brock, B. Herberich, P. G. Schultz, Science 2001, 292,
Ga-Ga-Cipso array which indicates lone pair character at the
gallium center,[7±11] 2) the NaÀaryl ring[8] and NaÀGa inter-
[2] L. Wang, A. Brock, P. G. Schultz, J. Am. Chem. Soc. 2002, 124, 1836 ±
actions[11] which shorten the GaÀGa distance, and 3) the role
[3] L. Wang, P. G. Schultz, Chem. Commun. 2002, 1 ± 11.
of the para-iPr groups on the flanking aryl rings which cause
[4] L. Wang, T. J. Magliery, D. R. Liu, P. G. Schultz, J. Am. Chem. Soc.
Ga-Ga-C angular distortions that can strengthen the GaÀGa
bond.[11] Force constant calculations have also pointed to a
[5] L. Wang, P. G. Schultz, Chem. Biol. 2001, 8, 883 ± 890.
relatively weak GaÀGa interaction.[12, 13] The publication of
[6] B. A. Steer, P. Schimmel, J. Biol. Chem. 1999, 274, 35 601 ± 35 606. [7] P. Brick, T. N. Bhat, D. M. Blow, J. Mol. Biol. 1989, 208, 83 ± 98.
the cluster species K2Ar*Ga4Ar*, which contains a Ga4 ring
[8] A second mutant M. jannaschii TyrRS library was also constructed in
with no GaÀGa triple bonding as part of an octahedral K2Ga4
such a way that six residues (Tyr 32, Ala 67, His 7 0, Gln 155, Asp 158,
core, has underlined the importance of the alkali metal for the
Ala 167) within 6.9 ä of the meta position of the tyrosine aryl ring
stability of 1.[14] However, apart from this isolated report, all
were randomly mutated. This library was also subjected to the genetic
arguments regarding the GaÀGa bonding in 1 have been
selection, but failed to produce a mutant TyrRS which charges O-allyl-l-tyrosine.
grounded in calculations of various degrees of sophistica-
[9] D. R. Liu, P. G. Schultz, Proc. Natl. Acad. Sci. USA 1999, 96, 4780 ±
tion[2±11] and the original experimentally determined structural
parameters.[1] In 1998 several experiments were suggested
[10] B. Nilsson, T. Moks, B. Jansson, L. Abrahmsen, A. Elmblad, E.
whose object was the elucidation of the important factors
Holmgren, C. Henrichson, T. A. Jones, M. Uhlen, Protein Eng. 1987, 1,107± 113.
governing the nature of the GaÀGa bond.[15] Among these
[11] We have also successfully incorporated 2 into mouse dihydrofolate
were the investigation of the effects of changing or removing
reductase (DHFR) with high fidelity (> 99 %).
the alkali metal ions and the isolation and characterization of
[12] The top two bands in Figure 1 with molecular weights of approx-
the neutral ™digallene∫ species Ar*GaGaAr* which, should
imately 32 kD and 23 kD are E. coli proteins which bind to the Ni2
contain a Ga-Ga double bond if the assumption of triple
[13] A. F¸rstner, Angew. Chem. 2000, 112, 3140 ± 3172; Angew. Chem. Int.
bonding in 1 was correct. The former question has been partly
answered through the synthesis of K2Ar*Ga4Ar*. However,
[14] K. V. Gothelf, K. A. Jorgensen, Chem. Rev. 1998, 98, 863 ± 909.
no stable neutral GaÀGa bonded dimers of the general
[15] S. C. G. Biagini, S. E. Gibson, S. P. Keen, J. Chem. Soc Perkin Trans. 1
formula RGaGaR (R organic or related group) have yet
[16] J. F. Reichwein, C. Versluis, R. M. J. Liskamp, J. Org. Chem. 2000, 65,
been described. Calculations on a variety of model species,
including HGaGaH,[10, 11, 16±19] MeGaGaMe,[9±11] and PhGa-
[17] H. E. Blackwell, J. D. Sadowsky, R. J. Howard, J. N. Sampson, J. A.
GaPh,[11] as well as IR spectroscopy of HGaGaH in a frozen
Chao, W. E. Steinmetz, D. J. O×Leary, R. H. Grubbs, J. Org. Chem.
matrix[19, 20] point to weak GaÀGa bonding. It is now shown
[18] D. M. Lynn, B. Mohr, R. H. Grubbs, L. M. Henling, M. W. Day, J. Am.
Dipp 2,6-iPr2C6H3), can be isolated and characterized with
[19] J. C. M. van Hest, K. L. Kiick, D. A. Tirrell, J. Am. Chem. Soc. 2000,
[20] K. L. Kiick, J. C. M. van Hest, D. A. Tirrell, Angew. Chem. 2000, 112,
2232 ± 2236; Angew. Chem. Int. Ed. 2000, 39, 2148 ± 2152.
[*] Prof. P. P. Power, Dr. N. J. Hardman, R. J. Wright, Dr. A. D. Phillips
[21] J. M. Fraile, J. I. Garcia, J. A. Mayoral, A. J. Royo, Tetrahedron:
University of CaliforniaOne Shields Avenue, Davis, CA 95616 (USA)Fax: ( 1) 530-752-8995E-mail: pppower@ucdavis.edu
[**] This work was supported by the National Science Foundation Grant
¹ WILEY-VCH Verlag GmbH, 69451 Weinheim, Germany, 2002
Tillæg til Cardiologisk Forum Oktober 2002 Nye retningslinjer vedrørende diagnostik og behandling Olav W.Nielsen, Niels Gadsbøll, Lars Køber, Per Hildebrandt, Henrik Villadsen Fra Arbejdsgruppen om Hjerteinsufficiens under Dansk Cardiologisk Selskab Introduktion 2. Objektiv dokumentation for kardial dysfunktionI 2001 udkom en task-force rapport om diagnostik3. Ved tvivl kan diagnosen
To be retuned at least 2 working days prior to appointment with nurse. Please complete this form prior to your travel appointment and return to reception. Personal Details Name: _______________________________________________________________________________________________ Date of birth: _____________________________________________________________________________________________

 [12] The side product related to 6 was also formed, but the optical purity
of O-allyl-l-tyrosine (2) into proteins in E. coli. The alkene
functional group of this unnatural amino acid should provide
[13] M. Shimazaki, H. Haram, K. Suzuki, G.-i. Tsuchihashi, Tetrahedron
new chemical methods for the selective modification of
Lett. 1987, 28, 5891; K. Suzuki, E. Katayama, G.-i. Tsuchihashi,
[14] Rearrangements of optically active vinyl epoxides mediated by Lewis
Previously we generated an orthogonal tRNA ± synthetase
acids give quaternary asymmetric carbon centers with inversion of the
pair, mutRNATyr ± TyrRS, in E. coli by modifying the
configuration: a) M. E. Jung, D. C. D×Amino, J. Am. Chem. Soc. 1995,
tRNATyr ± TyrRS pair of Methanococcus jannaschii.[4, 5] This
117, 7379; b) for rearrangements that involve a sterically restricted
mutRNATyr is not aminoacylated by endogenous synthetases
cation center, see: S. A. Monti, S.-C. Chen, Y.-L. Yang, S.-S. Yuan,
O. P. Bourgeois, J. Org. Chem. 1978, 43, 4062.
[12] The side product related to 6 was also formed, but the optical purity
of O-allyl-l-tyrosine (2) into proteins in E. coli. The alkene
functional group of this unnatural amino acid should provide
[13] M. Shimazaki, H. Haram, K. Suzuki, G.-i. Tsuchihashi, Tetrahedron
new chemical methods for the selective modification of
Lett. 1987, 28, 5891; K. Suzuki, E. Katayama, G.-i. Tsuchihashi,
[14] Rearrangements of optically active vinyl epoxides mediated by Lewis
Previously we generated an orthogonal tRNA ± synthetase
acids give quaternary asymmetric carbon centers with inversion of the
pair, mutRNATyr ± TyrRS, in E. coli by modifying the
configuration: a) M. E. Jung, D. C. D×Amino, J. Am. Chem. Soc. 1995,
tRNATyr ± TyrRS pair of Methanococcus jannaschii.[4, 5] This
117, 7379; b) for rearrangements that involve a sterically restricted
mutRNATyr is not aminoacylated by endogenous synthetases
cation center, see: S. A. Monti, S.-C. Chen, Y.-L. Yang, S.-S. Yuan,
O. P. Bourgeois, J. Org. Chem. 1978, 43, 4062.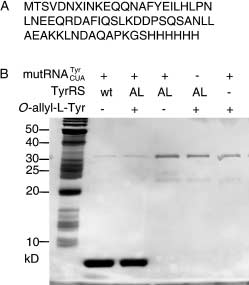 phenicol in the presence of 2, and up to 10 mg mLÀ1
chloramphenicol in the absence of 2. For comparison, E. coli
mutRNATyr ± AL-TyrRS was analyzed by electrospray ion-
cells expressing the Asp112amber CAT gene and the
ization Fourier transform ion cyclotron resonance mass
gene survive in 4 mg mLÀ1 chloramphenicol.[5]
spectrometry (FT-ICR MS). The experimental monoisotopic
This result suggests that the AL-TyrRS has higher activity for
mass of the intact protein was 7963.891 Da, which is within
2 than for natural amino acids. The evolved AL-TyrRS has the
1 ppm of the theoretical mass of 7963.889 Da. Another major
following mutations: Glu 107 3Ala 107, Asp 158 3Cys 158,
signal corresponds to the protein without the first methionine
and Ile 159 3Ala 159. The residues Tyr 32 and Leu 162 remain
unchanged. The mutations of Glu 107 3Ala 107and Ile 159
The signal-to-noise ratio of more than 1500 observed in the
3Ala 159 may enlarge the active site of the mutant synthetase
intact protein mass spectrum suggests a fidelity for the
to accommodate the allyl group. The exact roles of these
incorporation of 2 of better than 99.8 %. This result clearly
mutations will be revealed by solving the crystal structure of
demonstrates the site-specific incorporation of 2 in response
to the amber codon by the mutRNATyr ± AL-TyrRS, and that
To confirm that the observed phenotype is caused by the
other endogenous E. coli synthetases do not utilize 2 as a
site-specific incorporation of 2 by the mutRNATyr ± AL-
TyrRS pair, a mutant Z domain[10, 11] protein was produced
In summary, a useful nonnatural amino acid, O-allyl-l-
and characterized. An amber codon was introduced at the
tyrosine, has been site-specifically incorporated into proteins
seventh position in the gene encoding the Z domain. A His6
in vivo. The allyl group is versatile in organic transformations,
tag was added to the C terminus of the Z domain to facilitate
including metathesis,[13] Diels-Alder and 1,3-dipolar cyclo-
protein purification by Ni2-affinity chromatography. As a
addition reactions.[14] Olefin metathesis has been used suc-
positive control, the wild type M. jannaschii TyrRS was
cessfully to cross-link amino acid derivatives[15] and cyclize
coexpressed with the mutRNATyr , which resulted in the
peptides.[16, 17] Recent progress in developing water-soluble
suppression of the amber codon with tyrosine and production
ruthenium catalysts[18] should facilitate the application of this
of full-length Z-domain protein (Figure 1). When AL-TyrRS
reaction to the modification of proteins containing alkenegroups.[19, 20] Moreover, the allyl side chain of this nonnaturalamino acid may confer new physical properties on proteins aswell.
phenicol in the presence of 2, and up to 10 mg mLÀ1
chloramphenicol in the absence of 2. For comparison, E. coli
mutRNATyr ± AL-TyrRS was analyzed by electrospray ion-
cells expressing the Asp112amber CAT gene and the
ization Fourier transform ion cyclotron resonance mass
gene survive in 4 mg mLÀ1 chloramphenicol.[5]
spectrometry (FT-ICR MS). The experimental monoisotopic
This result suggests that the AL-TyrRS has higher activity for
mass of the intact protein was 7963.891 Da, which is within
2 than for natural amino acids. The evolved AL-TyrRS has the
1 ppm of the theoretical mass of 7963.889 Da. Another major
following mutations: Glu 107 3Ala 107, Asp 158 3Cys 158,
signal corresponds to the protein without the first methionine
and Ile 159 3Ala 159. The residues Tyr 32 and Leu 162 remain
unchanged. The mutations of Glu 107 3Ala 107and Ile 159
The signal-to-noise ratio of more than 1500 observed in the
3Ala 159 may enlarge the active site of the mutant synthetase
intact protein mass spectrum suggests a fidelity for the
to accommodate the allyl group. The exact roles of these
incorporation of 2 of better than 99.8 %. This result clearly
mutations will be revealed by solving the crystal structure of
demonstrates the site-specific incorporation of 2 in response
to the amber codon by the mutRNATyr ± AL-TyrRS, and that
To confirm that the observed phenotype is caused by the
other endogenous E. coli synthetases do not utilize 2 as a
site-specific incorporation of 2 by the mutRNATyr ± AL-
TyrRS pair, a mutant Z domain[10, 11] protein was produced
In summary, a useful nonnatural amino acid, O-allyl-l-
and characterized. An amber codon was introduced at the
tyrosine, has been site-specifically incorporated into proteins
seventh position in the gene encoding the Z domain. A His6
in vivo. The allyl group is versatile in organic transformations,
tag was added to the C terminus of the Z domain to facilitate
including metathesis,[13] Diels-Alder and 1,3-dipolar cyclo-
protein purification by Ni2-affinity chromatography. As a
addition reactions.[14] Olefin metathesis has been used suc-
positive control, the wild type M. jannaschii TyrRS was
cessfully to cross-link amino acid derivatives[15] and cyclize
coexpressed with the mutRNATyr , which resulted in the
peptides.[16, 17] Recent progress in developing water-soluble
suppression of the amber codon with tyrosine and production
ruthenium catalysts[18] should facilitate the application of this
of full-length Z-domain protein (Figure 1). When AL-TyrRS
reaction to the modification of proteins containing alkenegroups.[19, 20] Moreover, the allyl side chain of this nonnaturalamino acid may confer new physical properties on proteins aswell. Protein expression, purification, and characterization: Plasmid pLEIZ was
Synthesis andCharacterization of the Neutral
used to express the Z-domain gene with an amber codon at the seventh
position under the control of a bacteriophage T5 promoter and t
nator, and the mutRNATyr gene under the control of the lpp promoter and
Na2Ar'GaGaAr' (Ar' 2,6-Dipp2C6H3, Dipp
rrnC terminator. The AL-TyrRS gene was encoded in plasmid pBK-ALRS
under the control of the constitutive E. coli GlnRS promoter andterminator. E. coli DH10B cells cotransformed with pLEIZ and pBK-
Ned J. Hardman, Robert J. Wright, Andrew D. Phillips,
ALRS were grown in minimal media containing 1 % glycerol and 0.3 mm
leucine (GMML media) with 25 mg mLÀ1 kanamycin, 34 mg mLÀ1 ofchloramphenicol, and 0.5 mm 2. When cells reach an optical density
The publication of the remarkable gallium compound
(OD600) value of 0.5, isopropyl-b-d-thiogalactopyranoside (IPTG; 1 mm)was added to induce protein expression. After 4 h, cells were pelleted and
the protein was purified by Ni2-affinity chromatography according to the
iPr3C6H2) in 1997resulted in much controversy owing to the
manufacturer×s protocol under denaturing conditions (Quiagen, Valencia,
claim that it contained a GaÀGa triple bond.[1] Initially, the
CA). Proteins were then desalted by using a PD-10 column (Amersham
triple bonding in this molecule was justified on the basis of a
Pharmacia, Piscataway, NJ) and eluted in water. The yield of protein was
short GaÀGa distance (2.319(3) ä) and the correspondence of
measured by Bradford assay (BCA kit, Biorad, Hercules, CA). Aliquots ofprotein were used for SDS-PAGE and mass spectroscopic analysis.
Protein expression, purification, and characterization: Plasmid pLEIZ was
Synthesis andCharacterization of the Neutral
used to express the Z-domain gene with an amber codon at the seventh
position under the control of a bacteriophage T5 promoter and t
nator, and the mutRNATyr gene under the control of the lpp promoter and
Na2Ar'GaGaAr' (Ar' 2,6-Dipp2C6H3, Dipp
rrnC terminator. The AL-TyrRS gene was encoded in plasmid pBK-ALRS
under the control of the constitutive E. coli GlnRS promoter andterminator. E. coli DH10B cells cotransformed with pLEIZ and pBK-
Ned J. Hardman, Robert J. Wright, Andrew D. Phillips,
ALRS were grown in minimal media containing 1 % glycerol and 0.3 mm
leucine (GMML media) with 25 mg mLÀ1 kanamycin, 34 mg mLÀ1 ofchloramphenicol, and 0.5 mm 2. When cells reach an optical density
The publication of the remarkable gallium compound
(OD600) value of 0.5, isopropyl-b-d-thiogalactopyranoside (IPTG; 1 mm)was added to induce protein expression. After 4 h, cells were pelleted and
the protein was purified by Ni2-affinity chromatography according to the
iPr3C6H2) in 1997resulted in much controversy owing to the
manufacturer×s protocol under denaturing conditions (Quiagen, Valencia,
claim that it contained a GaÀGa triple bond.[1] Initially, the
CA). Proteins were then desalted by using a PD-10 column (Amersham
triple bonding in this molecule was justified on the basis of a
Pharmacia, Piscataway, NJ) and eluted in water. The yield of protein was
short GaÀGa distance (2.319(3) ä) and the correspondence of
measured by Bradford assay (BCA kit, Biorad, Hercules, CA). Aliquots ofprotein were used for SDS-PAGE and mass spectroscopic analysis.