Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Ber 5: 39-52
BOREAL ENVIRONMENT RESEARCH5: 39–52
This paper was presented at the symposium ‘Integrated Lake and Landscape Management’ (18–21 August1997, Lahti, Finland) under the auspices of the LIFE project ‘Integrated System of Drainage Area and WaterRehabilitation’ (FIN/A17/FIN/105/PIJ; coordinated by prof. T. Kairesalo)
Restoration of the eutrophicated Köyliönjärvi(SW Finland) through fish removal: whole-lakevs. mesocosm experiences
Jouko Sarvala1), Anne-Mari Ventelä1), Harri Helminen2),Arto Hirvonen3), Vesa Saarikari1), Seppo Salonen4), Asko Sydänoja1)and Kristiina Vuorio1)
1) Department of Biology, University of Turku, FIN-20014 Turku, Finland2) SW Finland Regional Environment Centre, Inkilänkatu 4, FIN-20300 Turku,
3) Köyliönjärvi Restoration Project, FIN-27710 Köyliö, Finland4) Satakunta Environmental Research Centre, University of Turku, Konttorikatu 1,
Sarvala, J., Ventelä, A.-M., Helminen, H., Hirvonen, A., Saarikari, V., Salonen, S.,Sydänoja, A. & Vuorio, K. 2000. Restoration of the eutrophicated Köyliönjärvi,southwestern Finland through fish removal: whole-lake vs. mesocosm experi-ences. Boreal Env. Res. 5: 39–52. ISSN 1239-6095
To improve water quality in a heavily eutrophicated lake (Köyliönjärvi, SW Finland),mass removal of fish was performed in 1992–1998. The fish stock declined from anestimated 170–250 kg ha–1 in 1991–1992 to 40–90 kg ha–1 in 1996–1998 or to 12%–25% of the initial biomass. The biomass of the larger cladocerans slightly increased in1991–1996 but decreased again in 1997, and chlorophyll a levels varied inversely withthe cladoceran biomass. Cyanobacteria initially declined, but altogether the water qual-ity effects of fish removal remained small up to the summer 1997. The roles of phos-phorus, submerged macrophytes (Elodea) and fish were further explored in a factorialenclosure experiment. Significant treatment effects were only observed in the earliestphase of the experiment, when the presence of macrophytes decreased and that of fishincreased phytoplankton chlorophyll a; later fish treatments were lost. Phosphorus ad-ditions had no effect on water quality, but at the end of the experiment phytoplanktonchlorophyll a was negatively correlated with the biomass of large cladocerans and posi-tively correlated with total phosphorus concentration. The enclosure effect was strong,all enclosures having much lower nutrient and chlorophyll levels than the surroundinglake. The experiment suggests that it is possible to improve water quality through re-moval fishing even in hypertrophic lakes, but the fish stock, including the young-of-the-year fish, must be decimated to a very low level. Introduction
stocking of piscivorous fish (young-of-the-yearand age 1+ pikeperch and 0+ pike; Salonen et al.
Increasing eutrophication is a common problem
1996, 1998). The removal of coarse fish has so
all over the world. Although the primary cause of
far continued from 1992 to 1998 (Hirvonen and
eutrophication is excessive external loading of
Salonen 1995, Salonen et al. 1996). In this paper,
nutrients, especially phosphorus, attempts to re-
we describe the results of the removal fishing and
verse the eutrophication process by curbing the
the development of water quality in the lake, com-
external load have often failed (Marsden 1989,
plementing the presentation of Sarvala et al.
Jeppesen et al. 1991). The dense populations of
(1998) with new data. We also examine the fu-
cyprinid fish in eutrophic lakes maintain a strong
ture prospects of the restoration project on the
internal loading/cycling of nutrients and control
herbivorous zooplankton, thus slowing down orpreventing the improvement of water quality. Re-cently, food web manipulation through removal
Study area
of excess planktivorous and benthivorous fish hasbecome a popular way to speed up the restoration
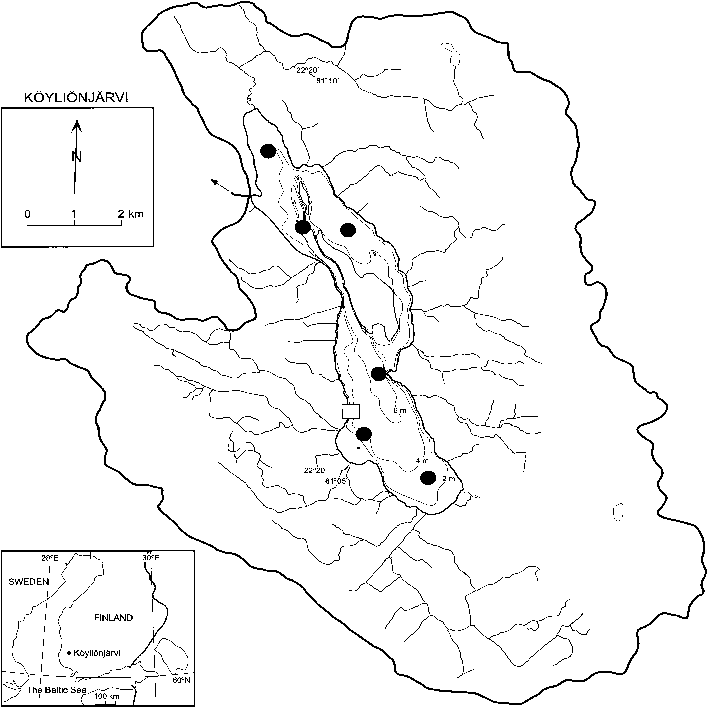
Köyliönjärvi (61°05´–61°10´N, 22°18´–22°24 ´E;
of eutrophicated lakes, usually in combination
Fig. 1) is shallow lake (mean depth 3.0 m, maxi-
with reductions in external load (Benndorf 1990,
mum depth 13 m), and therefore does not show
Reynolds 1994, Horppila et al. 1998).
any permanent temperature stratification during
Köyliönjärvi, a lake in southwestern Finland,
summer. The lake is normally ice-covered for 6
is an example of a culturally eutrophicated lake
months from early November to late April or early
amidst of an intensively cultivated agricultural
May. The drainage area is 129 km2 and the lake
area. It is shallow and located in an area of fertile
area is 12.5 km2. Theoretical water retention time
soils and has thus probably always been relatively
is 1.0 years. The lake is highly eutrophic, the late
productive. However, during the last decades it
has become hypertrophic mainly due to intensi-
reached up to 170 mg P m–3 and those of chloro-
fied agriculture (Itkonen and Olander 1997). To-
phyll a up to 180 mg chl a m–3 (Sarvala et al.
tal phosphorus levels in water typically increase
1998). Extensive cyanobacterial blooms have
during the summer and reach very high values in
occurred in late summer, and transparency is poor,
late summer (Sarvala et al. 1995). The late sum-
late summer Secchi depth being 0.3–0.5 m (Sar-
mer total phosphorus levels in water seem to have
vala et al. 1995). The external phosphorus load-
increased exponentially since the 1960s, or prob-
ing (0.64 g P m–2 a–1; Wright et al. 1993) exceeds
ably since the internal loading from the sediments
the “permissible” limits of Vollenweider (1975)
became important. Concomitant with the increas-
by a factor of five. About 93% of the total phos-
ing nutrient levels, phytoplankton biomass and
phorus input comes as diffuse loading from culti-
chlorophyll increased to hypertrophic levels, and
vated fields that comprise 32% of the drainage
heavy blooms of cyanobacteria became common.
area. Two thirds of the annual phosphorus load is
During the 1980s, the steadily deteriorating wa-
retained in the lake (Wright et al. 1993). There is
ter quality started to impede all uses of the lake,
no commercial fishery in the lake, and the local
and motivated the local community to search for
recreational and subsistence fishery utilizes
means to improve the situation: in 1990, a resto-
ration project was founded to rescue Köyliönjärvi. The long-term goal of the project was a notablereduction of nutrient loading from the surround-
Material and methods
ing agricultural area, but, in order to reach con-trol over the internal loading, and hoping to
Water quality and plankton
achieve more rapid progress, the project alsostarted a food web manipulation through removal
Nutrients, chlorophyll, phytoplankton and zoo-
fishing (Hirvonen et al. 1993, Sarvala et al. 1998).
plankton in Köyliönjärvi have been monitored
Later this was complemented with intensified
since 1991, mostly from weekly samples (twice a
Restoration of the eutrophicated Köyliönjärvi
Fig. 1. Bathymetric map of Köyliönjärvi, showing the sampling stations in the south and north basins (dots), as well as the site of the mesocosm experi- ment in 1996 (open square).
week in 1992). Two water columns from the sur-
cies had been measured. Length measurements
face to bottom were sampled with one-metre in-
were converted to carbon biomass using carbon
tervals with a 6.8-l tube sampler (Limnos Ltd.,
to length regressions (as in Sarvala et al. 1998).
Finland; in 1996 the sampler volume was 2.6 l
Additional water chemistry and chlorophyll
and in 1997 3.5 l) at each of three sites both in the
data were available from vertical sampling series,
southern and the northern basin of the lake (Fig. 1)
taken at 2–3 sites usually in late winter and late
and combined into a single composite sample for
summer since the 1960s (statutory monitoring and
each basin and date. Nutrient and chlorophyll
data obtained by the water authorities; unpublished
analyses were made in the laboratory of the South-
reports of the Water Protection Association of the
west Finland Regional Environment Centre, those
of phytoplankton and zooplankton in the Univer-sity of Turku. Phytoplankton samples (200 ml)were preserved with acid Lugol solution and
counted with an inverted microscope (Utermöhlsystem). Zooplankton samples were concentrated
Removal fishing was done in the winters 1992–
with a 25 or 50 mm mesh net and preserved with
1998 by commercial fishermen from the nearby
cold 94% ethanol (final concentration 70%). Us-
lake, Pyhäjärvi, (Sarvala et al. 1998) with seine
ing an inverted microscope, crustacean zooplank-
nets operated through holes in the ice. Because of
ton was identified and counted from subsamples
the bottom topography, the fishing concentrated
until 50–200 individuals of each dominant spe-
in the southern basin (area 400 ha). In 1996–1998,
seine nets were also used in open water in the
explored in 1996 in a 2 × 2 × 2 factorial enclosure
autumn, locating the aggregations of fish by echo
(6 m3) experiment with 3 replicates, following the
sounding and sonar. In 1992 and 1996, littoral trap
procedures used earlier in another lake, Vesijärvi
nets, operated by local inhabitants, were also used
(Kairesalo et al. 1998). The experimental enclo-
throughout the open-water season. Harvested fish
sures were attached to a wooden pier, built at a
were sold for animal feed production or directly
50 m distance from the western shore of the south
to fur farms. The average net cost of removal fish-
basin. The initial water depth at the site was 1.2–
ing was 2.50 FIM (0.42 ECU) kg–1 or about 150 FIM
1.4 m, and it declined during the experiment by
(25 ECU) ha–1 a–1 (Hirvonen et al. 1998).
20 cm. The pier comprised three contagious rows
Changes in the fish community of Köyliönjärvi
of eight 2 × 2.5 m frames parallel to the shore.
were studied from two-stage catch samples (pro-
The enclosures consisted of transparent 0.2-mm
cedure described in Salonen et al. 1996) taken
plastic, factory-moulded into wide tubes that were
from 322 (80.1%) of the total of 402 winter seine
attached within the frames. The enclosure walls
net hauls during 1992–1998. The samples cov-
were sealed into the sediment with sand bags (thir-
ered 94.3% of the total winter catch of 355 t. The
ty 2.5-kg bags per enclosure) inserted into a canal
composition of the trap net and open-water seine
welded along the lower ends of the walls. The
net catches (163 t) was similarly assessed. In1992–1993, the mesh size of the seine cod-end
underwater structures were checked by a diver.
was 8 mm, from 1994 onwards 6 mm. During the
The enclosures thus included both water and the
winter fishing seasons of 1992, 1993, 1996 and
underlying natural bottom sediment. Disturbances
1998, there was a significant decrease in the catch
by birds were prevented by covering the enclo-
per seine net haul in the southern basin, allowing
sure system with nets. Enclosures were closed on
estimates of the total catchable fish stock with the
12 June and checked by diving on 17 June.
removal (DeLury) method (for details of the meth-
od, see Hilborn and Walters 1992, and Helminen
domised block design, each row of enclosures
et al. 1993). To reduce random variation, three
containing all treatment combinations in random-
successsive hauls were combined for the final
ised order. For all factors, zero level denoted no
calculations, except in 1992 when only the single
additions. In the nutrient treatments, 70 mg P m–3
haul catches declined significantly. Stock esti-
as KH PO was added in the beginning and in the
mates for the southern basin were extrapolated to
middle of the experiment. In the macrophyte treat-
the whole lake using the ratio of surface areas.
ments, fresh Elodea collected from other parts of
Catch-effort estimates provided another way to
the lake were introduced in the beginning and
describe the fish stock development. We regressed
middle of the experiment. In fish enclosures, 16
the cumulative catch from the southern basin
individuals of 8–10 cm roach, caught with fyke
against the log-transformed number of seine hauls
nets from the lake, were introduced in the begin-
each winter. The smallest number of seine hauls
ning of the experiment. The first nutrient addi-
in any year from that basin was 26 hauls in 1997;
tions were done on 17 June, and the fish were
therefore, we calculated the expected catch after
introduced on 20 June. A total of 5.5 l of Elodea
26 hauls for the other years and used the actual
per treatment were added on three occasions (1 l
total catch for 1997. This method effectively
on 20 June, 2.5 l on 1 July and 2 l on 18 July; 1 l
smoothed the random variation of the catches; all
= 13.8 g dry mass). Sampling from the enclosures
regressions were highly significant. Age group
started on 23 June and ended 31 July. Very windy
analyses of roach (not presented here) allowed us
weather during the early half of the experiment
to compensate for the coarser mesh size used in1992–1993: the stock estimates for 1992 were
caused leakages of the wall plastic, and, conse-
increased by 15.5% and those for 1993 by 27.0%.
quently, the fish and first nutrient treatments werelost. After the leakages had been repaired (by 11July), the nutrient and macrophyte additions were
Mesocosm experiment
renewed, but no further fish were added. Nutri-ents, chlorophyll, bacteria, protozoa, phytoplank-
The roles of nutrients, macrophytes (Elodea) and
ton and zooplankton were sampled at 10-day in-
fish (8–10 cm roach (Rutilus rutilus (L.)) were
tervals (composite samples from the bottom and
Restoration of the eutrophicated Köyliönjärvi
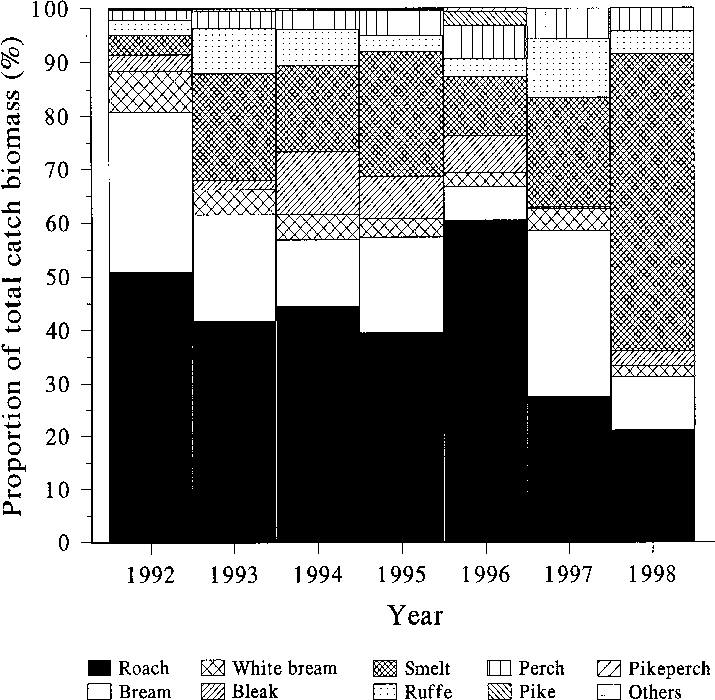
Fig. 2. Development of the total fish stock in Köyliön- järvi in 1991–1998. Dots denote DeLury estimates (ver- tical bars: 95% confidence limits); open squares de- note the cumulative catch per 26 hauls (vertical bars: standard error). Fig. 3. Proportions of different fish species in the re- moval fishing catch (winter seine only) in 1992–1998.
surface layers using a 0.5 m high tube sampler(Limnos; volume 3.5 l). Samples were also takenfrom the open lake in the immediate vicinity of
crease through the biomanipulation years (Fig. 2).
the enclosures. Periphyton development was as-
The DeLury estimates for the total fish stock had
sessed from plastic strips suspended vertically into
wide and variable confidence belts (Fig. 2). The
each enclosure (6 strips/enclosure), but no numeri-
point estimates were always close to the lower
cal results can be presented because the strips were
confidence bound, while the upper confidence belt
lost during storage. At the end of the experiment,
was much wider. According to DeLury estimates,
fish were removed by traps and handnets, and mac-
the fish biomass declined from roughly 170–250 kg
ha–1 in 1991–1992 to 40–90 kg ha–1 in 1996–1998
The experimental results were examined with
(Fig. 2). The catch-per-unit-effort figures sug-
factorial analysis of variance, multiple regression
gested a more regular and somewhat steeper de-
and partial correlation. The normality of variables
cline of the stock to about 12%–25% of the initial
was checked with the Wilks-Shapiro test. At the
end of the experiment, the values for chlorophyll,
Although there was some fluctuation in the
total phytoplankton biomass, cyanobacterial bio-
proportions of different species in the catch among
mass, total zooplankton biomass, total phospho-
the years (Fig. 3), the proportions of roach and
rus and total nitrogen did not deviate from nor-
bream seemed to decline somewhat (the share of
mal distributions, and the remaining variables
bream in 1997 was inflated by a single large indi-
(most of the individual zooplankton and phyto-
vidual in the catch samples), while the proportion
plankton groups) could be normalized with a log -
of smelt remained largely similar or increased,
and perch proportion slightly increased, exceptfor the last year. Altogether the proportion ofpiscivorous fish (pikeperch, pike and large perch
[> 10 cm]) remained very low (< 5%) throughoutthe period. There were some changes in the size
Removal fishing
distribution of the fish stock. Even correcting forthe effect of the larger mesh size in 1992–1993,
Altogether 518 tonnes (414 kg ha–1) of fish (mainly
the proportion of > 10 cm roach decreased during
roach and smelt [Osmerus eperlanus (L.)]) were
the period, especially because of strong year-
removed from Köyliönjärvi until the end of 1998.
classes hatched in 1996 and 1997 (Fig. 4); simul-
Both fish stock indices showed largely similar de-
taneously the mean age of roach declined from
Fig. 5. Average total phosphorus concentration in August in the south basin of Köyliönjärvi in 1966–1998. Sources: Water Protection Association of Kokemäenjo- ki Watercourse and Southwest Finland Regional En- vironment Centre.
but the between-year fluctuations, caused by e.g. weather differences, were too wide to allow defi-
Fig. 4. Length distribution of the roach caught with
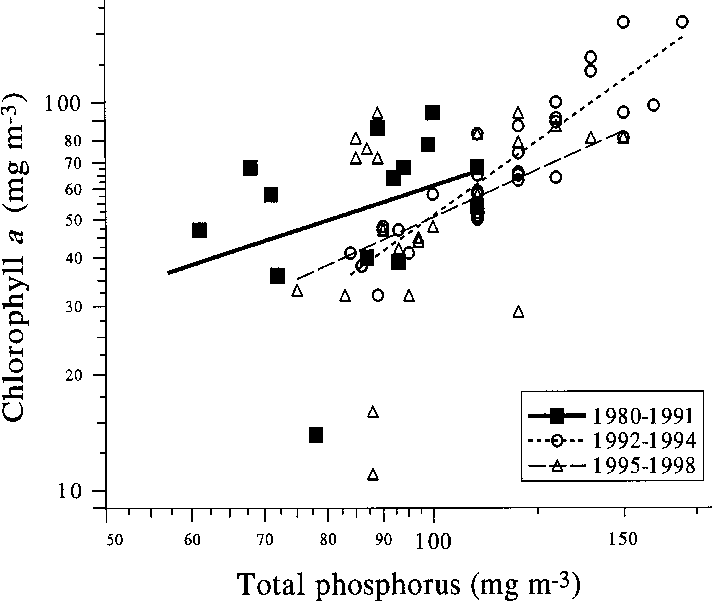
Altogether the water quality effects of fish
removal were so far small. During the biomanipu-
3.3 to 2.1 years (J. Sarvala unpubl. data). How-
lation period, chlorophyll levels relative to total
ever, the only species showing a significant trend
phosphorus in water were slightly lower than dur-
in size was bream, the mean size of which at least
ing the preceding decade (Fig. 9). In the early sum-
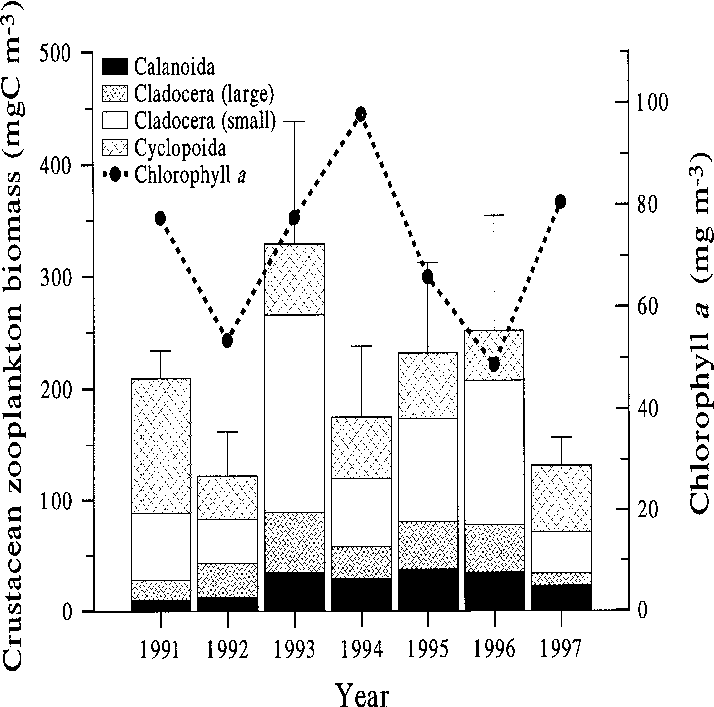
mers of 1996 and 1997, phytoplankton biomasswas clearly lower than in the previous years, andyet crustacean biomass remained high. The latesummer crustacean zooplankton biomass may
Water quality development
have slightly increased up to the year 1996, but in1997 there was a clear decline again (Fig. 7). Larg-
The late summer phosphorus concentrations in-
est between-year variation was due to small clado-
creased exponentially since the 1960s, but during
cerans, mainly Chydorussphaericus (Müller). In
the food web manipulation the values have started
both years, phytoplankton chlorophyll level and
to decrease (Fig. 5). The decreasing trend was most
the biomass of cyanobacteria increased again by
pronounced in late summer (Fig. 6). Late sum-
autumn. From the moderately decreased fish
mer chlorophyll levels did not show any consist-
biomass levels, larger water quality improvements
ent trend during the biomanipulation period, but
might have been expected in the summer 1997,
in the later years there was an inverse relationship
but the exceptionally high temperatures in that
with the herbivorous crustacean biomass (Fig. 7).
summer probably counteracted any positive de-
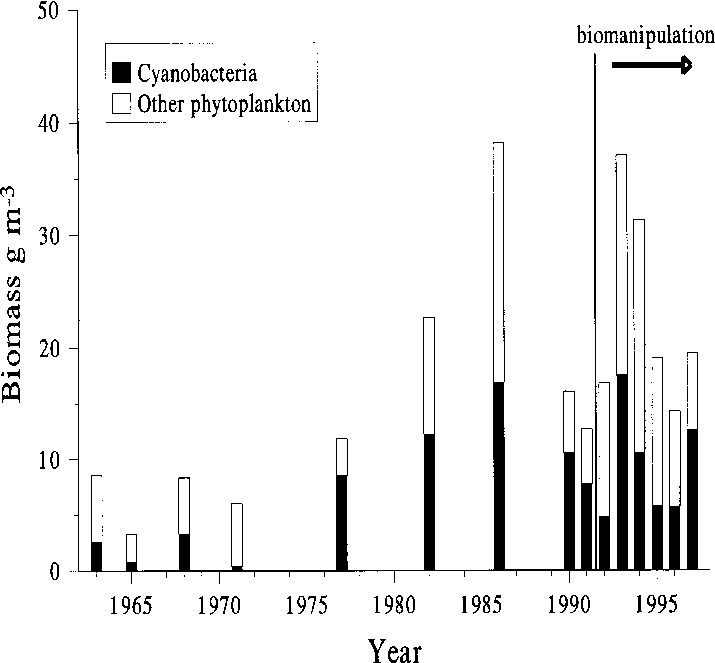
The total phytoplankton biomass increased
velopment (cyanobacterial blooms were then ex-
during the 1970s concomitant with the phospho-
tremely common in most watercourses in south-
rus concentrations, and simultaneously the pro-
ern Finland; unpublished data base of Finnish
portion of cyanobacteria increased, during the
Environment Institute). However, there were im-
1980s to 70%–80% of total phytoplankton bio-
portant changes within the cyanobacterial com-
mass in July–August (Fig. 8). There may be a
munity in summer 1997: the Microcystis species,
slight declining trend in the late summer total phy-
which had been dominant in previous years, were
toplankton biomass and in the contribution of
replaced by Anabaena. As a result, there were
cyanobacteria during the biomanipulation period,
almost no surface blooms of cyanobacteria in Köy-
Restoration of the eutrophicated Köyliönjärvi
Fig. 7. Late summer (26 July–15 September) concen- Fig. 6. Average total phosphorus concentrations in the
tration of chlorophyll a and the biomass of the main
south basin of Köyliönjärvi in May, June, July and Au-
crustacean zooplankton groups in the south basin of
gust in 1986–1998. Sources as in Fig. 5.
Köyliönjärvi in 1991–1997 (Sarvala et al. 1998, theyear 1997 added). Cladocera (“small”) denote Chydo-rus cf. sphaericus and Bosmina longirostris (Müller);Cladocera (“large”) include all other non-predatorycladoceran species. Fig. 8. The average biomass of total phytoplankton and the cyanobacteria in the south basin of Köyliönjärvi in July–August 1963–1997. Data for 1963–1990 from the Finnish Environment Institute.
liönjärvi in 1997 and the cyanobacterial toxins
Fig. 9. The average chlorophyll a concentration vs. total phosphorus in late summer in the south basin of
declined to a fraction of previous levels (J. Hietala,
Köyliönjärvi in 1980–1991 (before biomanipulation),
1992–1994 (early biomanipulation years) and 1995–1997 (late biomanipulation years). Data for 1980–1990from the Water Protection Association of Kokemäenjoki
Mesocosm experiment
Watercourse and Southwest Finland Regional Envi-ronment Centre.
Soon after the start of the experiment, water qual-ity in the enclosures began to diverge from the
showed widely divergent development. Differ-
surrounding lake (Fig. 10). This development was
ences were largest in chlorophyll and smallest in
reversed due to the leakages, and by 11 July whenthe leakages were repaired, most enclosures
nitrogen (Fig. 10). The relative homogeneity
showed phosphorus, nitrogen and chlorophyll val-
among the enclosures on 11 July, and the diver-
ues approaching those of the surrounding water.
gence on later dates, were also evident in the chlo-
During the next 20 days, different enclosures
rophyll: phosphorus relationship (Fig. 11). Total
Fig. 11. Chlorophyll a vs. phosphorus in different en- closures and in the surrounding lake during the meso- cosm experiment in Köyliönjärvi in 1996.
remained almost constant (range 58–90 mgP m–3),while most of the enclosures showed decreasingphosphorus levels. Total nitrogen in the lake in-creased throughout the period, but the relativeincrease was slightly less than in phosphorus. Inmost of the enclosures nitrogen remained morestable, but some showed similar increase as in thelake. In general, compared to the situation in phos-phorus or chlorophyll, the total nitrogen levels inthe enclosures followed more closely the condi-tions in the lake. In the lake, chlorophyll levelsincreased considerably in late July, but decreasedinstead in most enclosures.
At the end of the experiment, the total zoo-
plankton biomass was much higher in all enclo-sures than in the lake. The largest differences werein the biomass of herbivorous cladocerans (espe-cially Daphnia cucullata Sars and D. cristata Sars,sometimes also Ceriodaphnia pulchella Sars) andcalanoids (Eudiaptomus graciloides Lilljeborg). Chydorus sphaericus showed often the highestbiomass; there was negative correlation between
Fig. 10. Development of the total phosphorus, total
the Chydorus and Daphnia biomasses.
nitrogen and chlorophyll a concentrations in the 24
A significant effect of the fish (positive) and
experimental enclosures (macrophyte treatments
plant treatments (negative) on the chlorophyll
shown with thin solid lines) and the surrounding lake
level of water was observable on the first sam-
(thick lines with dots) during a mesocosm experiment
pling occasion when the enclosures were still in-
in Köyliönjärvi in summer 1996. Vertical arrows show
tact (23 June; ANOVA, Table 1; neither the block
effect nor any of the interactions were significantand their sums of squares were therefore pooled
phosphorus in the open lake increased mainly
into the error term). During the rest of the experi-
during early stages of the experiment and later
ment, the fish treatments must be ignored because
Restoration of the eutrophicated Köyliönjärvi
most fish escaped through the leakages and no
0.79) and chlorophyll and oxygen concentration
further fish were added. Plant and nutrient treat-
of water (r = 0.82) were likewise highly signifi-
ments were valid after 11 July, but they did not
cant, confirming that chlorophyll level was also a
then affect water quality (chlorophyll or nutrient
good indicator of primary productivity in the en-
concentrations) to any significant extent. Although
closures. The chlorophyll a concentrations in the
the added amount of phosphorus was theoretically
enclosures showed significant negative correla-
sufficient to double the total phosphorus concen-
tions with the biomasses of total crustaceans and
tration in the enclosures, no significant effects
the large herbivorous crustaceans (the larger
could be traced in any of the analyses. Elodea was
cladocerans Daphnia, Ceriodaphnia, Diaphano-
able to increase appreciably in only one enclo-
soma, Sida, Limnosida plus the calanoid Eudiapto-
sure; in most enclosures the plant biomass de-
mus graciloides), and significant positive corre-
creased. In contrast, the enclosure effect was
lations with the total phosphorus and nitrogen con-
strong, most enclosures having lower phospho-
centrations (Table 2). Among the negative corre-
rus, nitrogen and chlorophyll levels than the sur-
lations of chlorophyll with the major zooplankton
rounding lake. This was likely due to the thick
groups, those with the calanoid and, unexpectedly,
mat of periphyton that rapidly developed on the
cyclopoid biomass were significant. The latter cor-
plastic walls; changed sediment-water interactions
relation either indicates herbivorous feeding by
in the enclosures may also have been involved.
cyclopoids or the effect of a confounding factor.
However, the difference between the lake and the
Simple correlations between total nitrogen and
enclosures was much larger in chlorophyll than
large herbivorous crustaceans as well as between
in phosphorus or nitrogen, showing that factorsother than nutrients were also involved. This was
Table 1. Analysis of variance table for chlorophyll a in
also shown by the notable changes of chlorophyll
the enclosure experiment on 23 June 1996. DF = de-
at a certain phosphorus level (Fig. 11); changes
grees of freedom, SS = sum of squares, MS = mean
At the end of the mesocosm experiment, phy-
————————————————————————
toplankton biomass in the lake and in most enclo-
sures was dominated by cyanobacteria. Enclosures
————————————————————————
with low total biomass also had low proportion of
cyanobacteria. Chlorophyll a concentration was
a good measure of phytoplankton abundance, be-
ing tightly correlated with both total biomass (r =
0.91) and cyanobacterial biomass (r = 0.88). Posi-
tive correlations between chlorophyll and pH (r =
————————————————————————
Table 2. Simple correlations between phytoplankton chlorophyll a and various zooplankton and nutrient vari- ables at the end of the enclosure experiment (31 July 1996). Log -transformation used throughout (* = P < 0.05;
** = P < 0.01; N = 24). —————————————————————————————————————————————————
—————————————————————————————————————————————————Chlorophyll a (CHL)
Total crustacean zooplankton biomass (Z) –0.42*
—————————————————————————————————————————————————
feeding preferences of the crustacean groups.
Multiple regression analyses showed that phy-
toplankton chlorophyll a at the end of the experi-ment was predictable from the biomass of totalcrustacean zooplankton, large herbivorous crus-taceans, and the total phosphorus and total nitro-gen concentrations (Table 4, Fig. 13). The impor-tance of each nutrient in these regressions variedaccording to the other variables included. Multi-collinearity arising from the high mutual correla-tions between the independent variables makes itdifficult to judge the relative role of each explana-tory variable. When the effect of the other two
Fig. 12. Chlorophyll a vs. the third quartile of cladoce-
independent variables was removed, the highest
ran length distribution at the end of the mesocosm
partial correlation with chlorophyll was shown by
experiment in Köyliönjärvi in 1996. Triangle = lake
total crustacean zooplankton biomass (–0.54; large
herbivores: –0.42), and total phosphorus showedhigher partial correlation (0.47) than total nitro-
both phosphorus and nitrogen and the median or
gen (0.40) (ln-transformed values). Among the
third quartile of the cladoceran length distribu-
different zooplankton variables, the large herbiv-
tion were significant (Table 2, Fig. 12).
ore biomass explained the largest fraction (30%)
The relationships between different zooplank-
of chlorophyll variation. The correlations between
ton and phytoplankton groups showed interesting
chlorophyll and the median or third quartile of
variation (Table 3). The large herbivores Daphnia
the cladoceran zooplankton length distribution
and Eudiaptomus as well as cyclopoids all showed
(Table 2) disappeared when nutrient variables
similar negative correlations (although not all
were included. The final concentration of nitro-
exceeding the 0.05 significance level) with cyano-
gen in the enclosures was also inversely corre-
bacteria and diatoms, Daphnia also with chloro-
lated with the biomass of large herbivores. These
phytes. Bosmina likewise had a negative correla-
results suggest that zooplankton grazing control-
tion with chlorophytes, while the biomass of Chy-
led phytoplankton abundance and that phospho-
dorus was negatively correlated with chrysophy-
rus was the primary nutrient affecting phytoplank-
tes. There was an almost significant positive corre-
ton abundance, the correlation between chloro-
lation between Daphnia and cryptophytes, also
phyll and nitrogen arising secondarily, because
manifest at the composite “large herbivore” group
increases in nitrogen-fixing blue-green algae si-
level. These correlations might reflect differing
multaneously lead to elevated total nitrogen. Table 3. Simple correlations between major crustacean zooplankton groups and the main phytoplankton groups at the end of the enclosure experiment (31 July 1996). Log -transformation used for all variables except
cyanobacterial biomass. Phytoplankton groups: Cyano = Cyanobacteria, Crypto = Cryptophyceae, Chryso =Chrysophyceae, Diatomo = Diatomophyceae, Chloro = Chlorophyceae. The zooplankton group “Other Cladocera”mainly consists of Chydorus. —————————————————————————————————————————————————
—————————————————————————————————————————————————Bosmina
—————————————————————————————————————————————————
Restoration of the eutrophicated Köyliönjärvi
Fig. 13. Chlorophyll a vs. total phosphorus and crus- tacean zooplankton bio- mass at the end of the me- socosm experiment in Köyliönjärvi in 1996. Dia- mond = lake outside enclo- sures. Discussion
although removal fishing had reduced the totalfish biomass to 12%–25% of the initial level. How-
Our long-term study indicated only slight changes
ever, a mesocosm experiment showed that abun-
in the water quality of eutrophic Köyliönjärvi,
dant herbivore crustacean zooplankton had the
Table 4. Regressions of phytoplankton chlorophyll a on the zooplankton biomass, total phosphorus concentra- tion and total nitrogen concentration (log -transformation used throughout) at the end of the mesocosm experi-
ment in Köyliönjärvi in 1996. (1) Total crustacean zooplankton biomass used, (2) herbivore crustacean biomassused, (3) same as (2), but without nitrogen. SD = standard deviation; N = 24. —————————————————————————————————————————————————Variables
—————————————————————————————————————————————————(1)Total crustaceanzooplankton
(2)Large herbivorouscrustacean zooplankton
(3)Large herbivorouscrustacean zooplankton
—————————————————————————————————————————————————
potential to control phytoplankton development
experiment was that the enclosure effects were so
dominant over other factors affecting water qual-
Scheffer (1990) and Scheffer et al. (1993)
ity. The plastic enclosure walls provided a suit-
hypothesized that shallow lake ecosystems pos-
able firm surface for prolific growth of attached
sess two alternative stable states. At low nutrient
algae, which were then able to assimilate a nota-
levels submerged macrophytes abound and water
ble part of the nutrients in water into periphyton
is clear, while at high nutrient concentrations
biomass. Although rarely indicated in published
macrophytes disappear and water remains turbid.
reports, such strong enclosure effects can be ex-
Over a range of intermediate nutrient concentra-
pected to be common in comparable experiments.
tions the system might switch between these states
A negative correlation between periphyton and
depending on its history and recent perturbations.
phytoplankton has often been documented (Sand-
Our experiences from entire Köyliönjärvi confirm
Jensen and Borum 1991, Harris 1995). It is con-
the strong resistance to change of turbid, highly
ceivable that part of the macrophyte effects in such
eutrophic lakes (Jeppesen et al. 1991). Submerged
experiments might be mediated through epiphytic
macrophytes were not able to grow well in the
growth on the submerged plants (Van Donk et al.
turbid water neither in the enclosures nor in the
1995, Brock et al. 1995). In any case, the peri-
open lake, and thus they could not affect water
phyton development should be taken into account
quality. However, data collected so far from a
when interpreting experimental results.
number of southern Finnish lakes (Sarvala et al.
In contrast to several earlier studies (e.g.
1997, 1998, and unpublished) suggest that lakes
Schriver et al. 1995, Kairesalo et al. 1998), in our
may switch between turbid and clear state irre-
experiment the submerged macrophytes had only
spective of their nutrient levels. This result agrees
minor water quality effects. This was probably
well with the literature survey of Mazumder
due to the high turbidity of water that prevented
(1994) showing that the water quality responses
the growth of Elodea through light limitation. In
of lakes formed a wide belt over a range of nutri-
the experiments of Kairesalo et al. (1998) in
ent concentrations. For example, during a period
Vesijärvi, the initial transparency of water was
of 17 years, phytoplankton chlorophyll in Pyhäjär-
much better than in our experiment in Köyliönjär-
vi, a lake in southwestern Finland, did not oscil-
vi. Thus, although macrophytes clearly have the
late between two alternative end states, but, de-
potential to enhance water quality, this will not
pending on the strength of the planktivorous fish
come into effect until the transparency has first
stock, varied in a continuum between extremely
improved and remained good for a relatively long
high and extremely low values relative to the phos-
time to allow for macrophyte development (Irvine
phorus level (Sarvala et al. 1997, 1998) . Nothing
et al. 1989, Jeppesen et al. 1990).
in the behaviour of Pyhäjärvi suggests that any of
Although our mesocosm experiment was tech-
the annual situations would be an equilibrium
nically only partially successful, it suggested that
state, rather, the food web structure was continu-
the phytoplankton biomass was regulated not only
ously changing as a response to external (natural
by nutrients (phosphorus) but also to an almost
and anthropogenic) fluctuations; the nature of the
similar extent by the abundance of herbivorous
responses is affected by the different characteris-
crustacean zooplankton. The resulting multiple
tic time scales of the system components, e.g. due
regression equation was almost identical with the
to their long generation times, fish buffer changes
corresponding equation obtained from a long-term
and cause delayed responses. At the microbial
field data series for the neighbouring Pyhäjärvi
level the responses become evident within a cou-
(Sarvala et al. 1998), which is only weakly meso-
ple of days (e.g. A.-M. Ventelä unpubl.), while a
trophic. It seems therefore justified to conclude
strong year-class of fish may dominate the sys-
that probably the same mechanisms are regulat-
tem over several years. Like Persson et al. (1993),
ing water quality in both lakes, although compli-
we are sceptic about the existence of any real al-
cations may arise from the fact that the crusta-
ternative stable states in lake ecosystems.
cean zooplankton abundance also affected in sev-
eral ways the microbial food web in the enclo-
Restoration of the eutrophicated Köyliönjärvi
sures (K. Wiackowski unpubl.). Thus, although
References
the fish removal has not yet resulted in any dra-matic water quality changes in Köyliönjärvi, such
Benndorf J. 1990. Conditions for effective biomanipulation:
changes are likely when the crustacean zoo-
conclusions derived from whole-lake experiments in
plankton becomes more abundant. That crusta-
Europe. Hydrobiologia 200/201: 187–203.
Brock T.C.M., Roijackers R.M.M., Rollon R., Bransen F.
cean zooplankton has not yet increased as much
& Van der Heyden L. 1995. Effects of nutrient loading
as one might have expected from the decimated
and insecticide application on the ecology of Elodea-
fish populations, can most probably be attributed
dominated freshwater microcosms. II. Responses of
to two conditions. First, the remaining fish
macrophytes, periphyton and macroinvertebrate graz-
biomass consists mostly of very small-sized fish,
ers. Arch. Hydrobiol. 134: 53–74.
which are the most efficient plankton-feeders.
Chadwick E.M.P. 1976. Ecological fish production in a small
Precambrian shield lake. Env. Biol. Fish. 1: 13–60.
Second, exceptionally large year-classes of roach
Cryer M.G., Peirson G. & Townsend C.R. 1986. Recipro-
and other coarse fish were probably born during
cal interactions between roach (Rutilus rutilus L.) and
the very warm summer of 1997. It is known that
zooplankton in a small lake: prey dynamics and fish
the young-of-the-year fish often account for a
growth and recruitment. Limnol. Oceanogr. 31: 1022–
major part of total fish production (Chadwick
1976) and food consumption (Helminen et al.
Harris P.M. 1995. Are autecologically similar species also
functionally similar? A test in pond communities. Ecol-
1990, Helminen and Sarvala 1994), and, conse-
quently, are also most important in controlling the
Helminen H. & Sarvala J. 1994. Changes in zooplanktivory
crustacean zooplankton (Langeland 1982, Le-
by vendace (Coregonus albula) in Lake Pyhäjärvi (SW
scher-Moutoué et al. 1985, Cryer et al. 1986,
Finland) due to variable recruitment. Verh. Int. Ver.
Whiteside 1988, Helminen et al. 1990, Helminen
Helminen H., Sarvala J. & Hirvonen A. 1990. Growth and
food consumption of vendace (Coregonus albula (L.))
Based on our results, we conclude that it is
in Lake Pyhäjärvi, SW Finland: a bioenergetics model-
possible to improve water quality through removal
ing analysis. Hydrobiologia 200/201: 511–522.
fishing even in hypertrophic lakes, but the plankti-
Helminen H., Ennola K., Hirvonen A. & Sarvala J. 1993.
vorous fish stock must be reduced to a very low
Fish stock assessment in lakes based on mass removal.
level, and the piscivorous fish stocks should si-
multaneously be enhanced to decimate the num-
Hilborn R.J. & Walters C.J. 1992. Quantitative fisheriesstock assessment. Choice, dynamics and uncertainty,
bers of fish that are too small to be controlled by
Hirvonen A. & Salonen S. 1995. Ravintoketjukunnostuksen
alkutaival Köyliönjärvellä [The first stage in restoring
Acknowledgements: This paper is part of a series of studies
Lake Köyliönjärvi by fish removal]. Vesitalous 36(3):
financed by the Academy of Finland (grants 1071292, 4158,
11–14. Abstract p. 39. [In Finnish with English abstract.]
31365, 35425 and 35619 to JS). We thank Ilkka Blomqvist,
Hirvonen A., Helminen H., Saarikari V., Salonen S., Vuorio
Hannu Koponen, Marianne Moilanen, Marjo Tarvainen,
K. & Sarvala J. 1993. Effects of cyprinid reduction on
Krzysztof Wiackowski and Olli Ylönen, the laboratory per-
water quality in Lake Köyliönjärvi, SW Finland. In
sonnel of Southwest Finland Regional Environment Cen-
Giussani G. & Callieri C. (eds.), Proc. 5th Internat.
tre, Satakunta Environmental Research Centre, and Hannu
conference on the conservation and management of
Uusi-Laurila, the manager of the Räpi Experimental Farm
lakes “Strategies for lake ecosystems beyond 2000”,
in Köyliö, who all helped in various ways with the field and
17–21 May, 1993 Stresa, Italy, Pallanza, pp. 113–116.
Hirvonen A., Helminen H., Salonen S., Sydänoja A. &
laboratory work. Lake Köyliönjärvi Protection Association
Sarvala J. 1998. Köyliönjärven ravintoketjukunnostuk-
and an Engineering Unit from the Finnish Army helped with
sessa on alkanut loppusuora. Suomen Kalastuslehti
the construction of the field enclosures. Dr. Arto Itkonen,
Department of Geology, University of Turku, kindly pro-
Horppila J., Peltonen H., Malinen T., Luokkanen E. &
vided the map template for Fig. 1. The removal fishing was
Kairesalo T. 1998. Top-down or bottom-up effects by
financed by the municipality and fisheries district of Köyliö,
fish: issues of concern in biomanipulation of lakes.
Köyliö Fish Farm Ltd, the Turku Fisheries District, South-
Restoration Ecology 6: 20–28.
west Finland Regional Environment Centre, Lake Köyliön-
Irvine K., Moss B. & Balls H. 1989. The loss of submerged
järvi Protection Association and the local farmer and vil-
plants with eutrophication. II. Relationships between
fish and zooplankton in a set of experimental ponds,
and conclusions. Freshwat. Biol. 22: 89–107. economic and management aspects, FAO & Fishing
Itkonen A. & Olander H. 1997. The origin of the hyper-
trophic state of a shallow boreal shield lake. Boreal
Sand-Jensen K. & Borum J. 1991. Interactions among phyto-
plankton, periphyton, and macrophytes in temperate
Jeppesen E., Kristensen P., Jensen J. P., Søndergaard M.,
freshwaters and estuaries. Aquatic Botany 41: 137–175.
Mortensen E. & Lauridsen T. 1991. Recovery resilience
Sarvala J., Helminen H. & Kirkkala T. 1997. Pyhäjärven
following a reduction in external phosphorus loading
veden laatu ja sitä säätelevät tekijät [Quality of the water
of shallow, eutrophic Danish lakes: duration, regulat-
in Lake Pyhäjärvi and factors controlling it]. Vesitalous
ing factors and methods for overcoming resilience.
38(3): 15–20, 33. [In Finnish with English abstract.]
Mem. Ist. Ital. Idrobiol. 48: 127–148.
Sarvala J., Helminen H., Hirvonen A. & Salonen S. 1995.
Jeppesen E., Søndergaard M., Mortensen E., Kristensen P.,
Köyliönjärven veden laatu ja siihen vaikuttavat
Riemann B., Jensen H. J., Müller J. P., Sortkjær O.,
mekanismit [Effects of cyprinid fish reduction on wa-
Jensen J. P., Christoffersen K., Bosselmann S. & Dall
ter quality of Lake Köyliönjärvi]. Vesitalous 36: 15–
P. 1990. Fish manipulation as a lake restoration tool in
17. Abstract p. 39. [In Finnish with English abstract.]
shallow, eutrophic temperate lakes 1: cross-analysis of
Sarvala J., Helminen H., Ripatti J.-P., Pruuki V. & Ruuhi-
three Danish case studies. Hydrobiologia 200/201: 205–
järvi J. 1992. Tehokalastuksen vaikutus Evon Syrjänalu-
sen särkikantaan [Impact of intensive fishing on the
Kairesalo T., Tátrai I. & Luokkanen E. 1998. Impacts of
roach population in Lake Syrjänalunen, southern Fin-
waterweed (Elodea canadensis Mich.) on fish-plank-
land]. Suomen Kalatalous 60: 191–205. [In Finnish with
ton interactions in the lake littoral. Verh. Int. Ver.
Sarvala J., Helminen H., Saarikari V., Salonen S. & Vuorio K.
Langeland A. 1982. Interactions between zooplankton and
1998. Relations between planktivorous fish abundance,
fish in a fertilized lake. Holarctic Ecology 5: 273–310.
zooplankton and phytoplankton in three lakes of dif-
Lescher-Moutoué F., Garnier J. & Pourriot R. 1985. Inter-
fering productivity. Hydrobiologia 363: 81–95.
actions entre les peuplements planctoniques et piscicole
Scheffer M. 1990. Multiplicity of stable states in freshwa-
du lac de Créteil: impact d’une reproduction exception-
ter systems. Hydrobiologia 200/201: 475–486.
nelle de Percidés. Bull. Ecol. 16: 9–17.
Scheffer M., Hosper S.H., Meijer M.-L., Moss B. & Jeppe-
Marsden M.W. 1989. Lake restoration by reducing exter-
sen E. 1993. Alternative equilibria in shallow lakes.
nal phosphorus loading: the influence of sediment phos-
Trends Ecol. Evol. 8: 275–279.
phorus release. Freshwat. Biol. 21: 139–162.
Schriver P., Bøgestrand J., Jeppesen E. & Søndergaard M.
Mazumder A. 1994. Phosphorus-chlorophyll relationships
1995. Impact of submerged macrophytes on fish-
under contrasting herbivory and thermal stratification:
zooplankton-phytoplankton interactions: large-scale en-
predictions and patterns. Can. J. Fish. Aquat. Sci. 51:
closure experiments in a shallow eutrophic lake. Freshwat. Biol. 33: 255–270.
Persson L., Johansson L., Andersson G., Diehl S. & Hamrin
Van Donk E., Prins H., Voogd H.M., Crum S.J.H. & Brock
S.F. 1993. Density dependent interactions in lake eco-
T.C.M. 1995. Effects of nutrient loading and insecti-
systems: whole lake perturbation experiments. Oikos
cide application on the ecology of Elodea-dominated
freshwater microcosms. I. Responses of plankton and
Reynolds C.S. 1994. The ecological basis for the success-
zooplanktivorous insects. Arch. Hydrobiol. 133: 417–
ful biomanipulation of aquatic communities. Arch.
Vollenweider R.A. 1975. Input-output models, with spe-
Salonen S., Helminen H. & Sarvala J. 1996. Feasibility of
cial reference to the phosphorus loading concept in lim-
controlling coarse fish populations through pikeperch
nology. Schweiz. Zeitschr. Hydrol. 37: 53–84.
(Stizostedion lucioperca) stocking in Lake Köyliönjärvi,
Whiteside M.C. 1988. 0+ fish as major factors affecting
SW Finland. Ann. Zool. Fennici 33: 451–457.
abundance pattern of littoral zooplankton. Verh. Int.
Salonen S., Helminen H. & Sarvala J. 1998. Compatibility
Ver. Limnol. 21: 1710–1714.
of recreational fisheries and ecological lake restoration
Wright J., Helminen H. & Hirvonen A. 1993. Köyliönjärven
in pikeperch (Stizostedion lucioperca L.) management
kujanjuoksu: ulkoinen fosforikuormitus [The gauntlet
in Lake Köyliönjärvi, SW Finland. In Hickley P. &
of Lake Köyliönjärvi: external phosphorus load]. Vesita-
Tompkins H. (eds.), Recreational fisheries — Social,lous 34(5): 29–33. [In Finnish with English abstract.]
Received 29 June 1999, accepted 8 November 1999
LA LIBERTÉ MARDI 9 JUIN 2009 MARDI-IMMO RADIO-TV MAGAZINE JARDINAGE Vivre sa sexualité,malgré le handicap SOCIÉTÉ • Les premiers «assistants sexuels» de Romandie reçoivent leur diplôme samedi. Quatre femmes et six hommes formés durant un an pour offrir sensualité, intimité et sexualité à des personnes handicapées. ANNICK MONOD garantissant l’intimité.
Welcome to our office. Our doctors and staff look forward to providing you with quality dental care in a friendly and relaxed atmosphere. Your initial visit will include review of your medical and dental history, taking necessary x-rays and an evaluation of your mouth. Our treatment recommendations are based on your health needs. Thorough care is our foremost consideration and compris
 Restoration of the eutrophicated Köyliönjärvi
Fig. 1. Bathymetric map of
Restoration of the eutrophicated Köyliönjärvi
Fig. 1. Bathymetric map of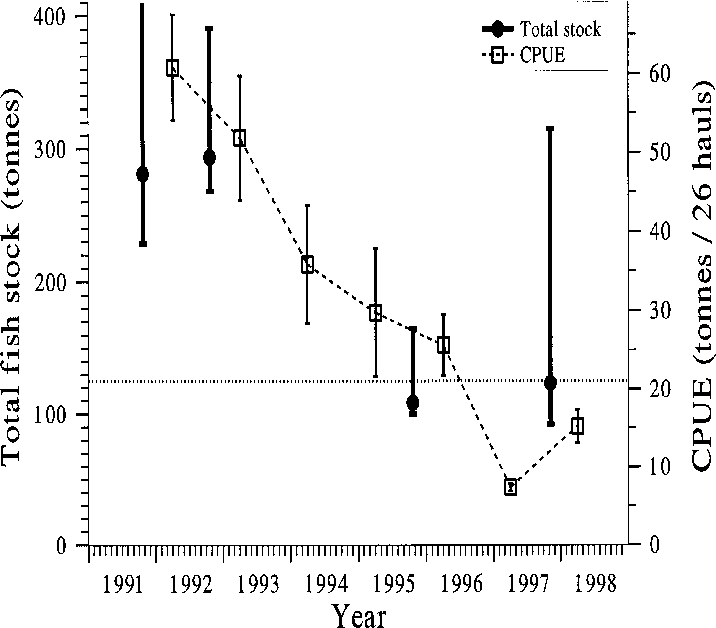
 Restoration of the eutrophicated Köyliönjärvi
Fig. 2. Development of the total fish stock in Köyliön-
Restoration of the eutrophicated Köyliönjärvi
Fig. 2. Development of the total fish stock in Köyliön-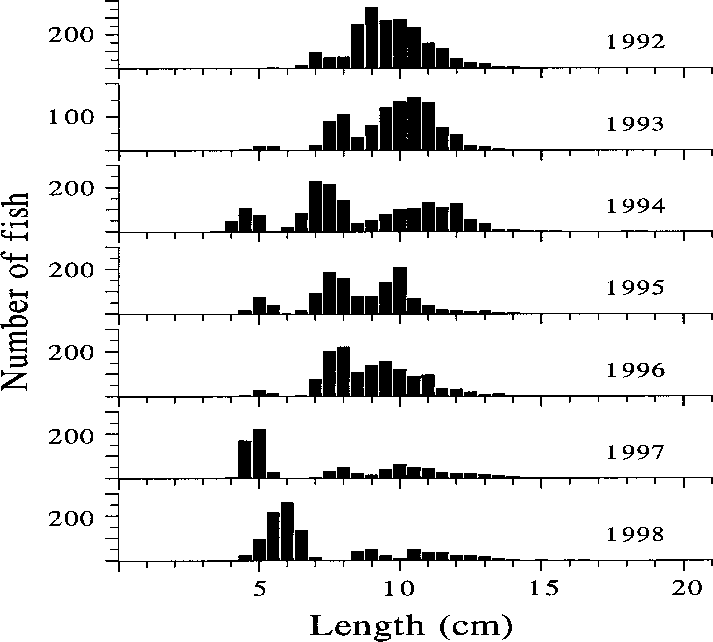
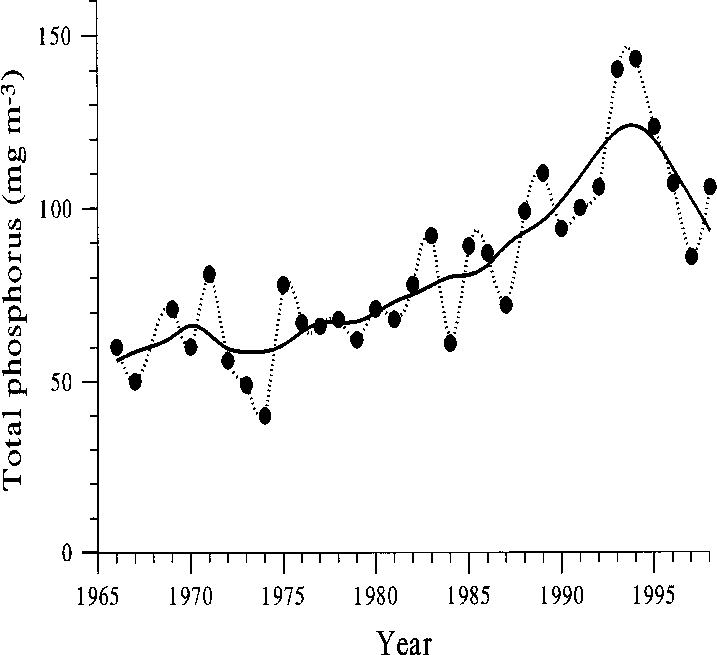 Fig. 5. Average total phosphorus concentration in
Fig. 5. Average total phosphorus concentration in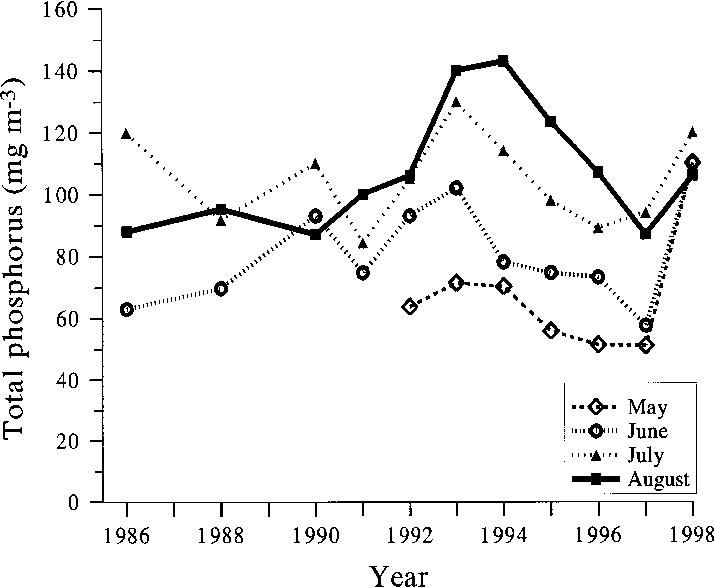
 Restoration of the eutrophicated Köyliönjärvi
Fig. 7. Late summer (26 July–15 September) concen-
Restoration of the eutrophicated Köyliönjärvi
Fig. 7. Late summer (26 July–15 September) concen-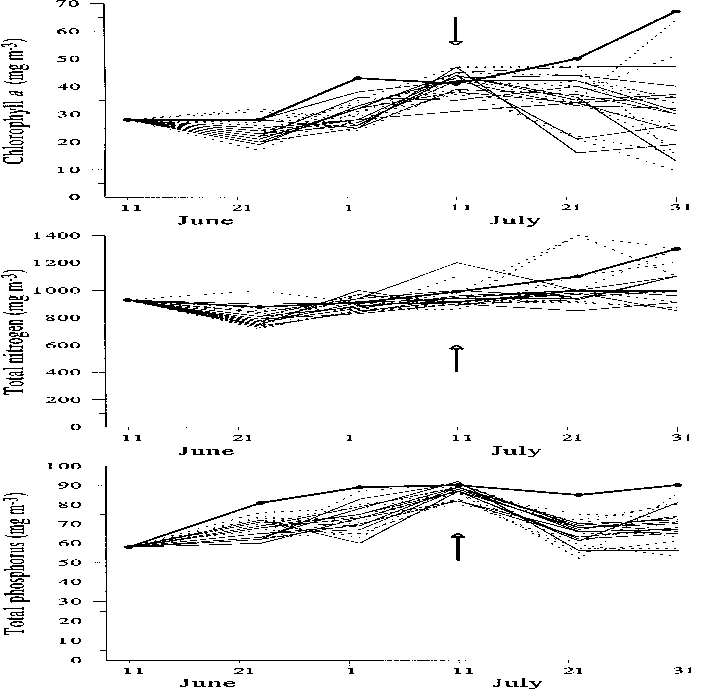
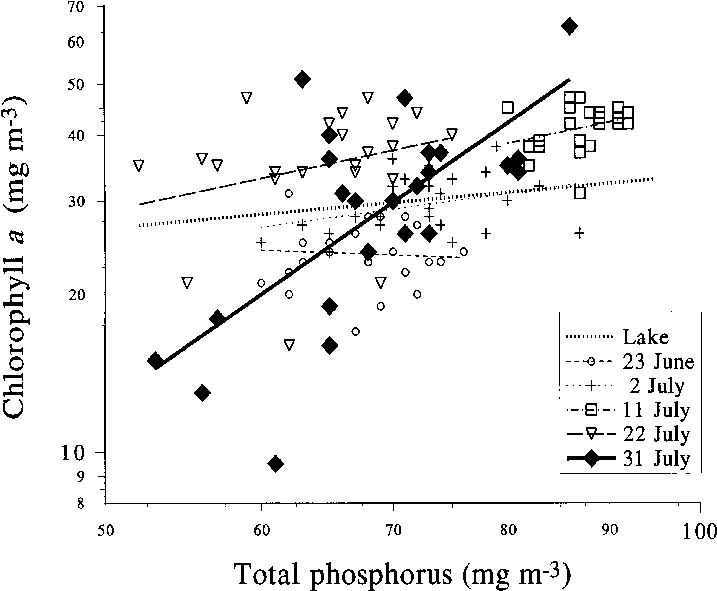 Fig. 11. Chlorophyll a vs. phosphorus in different en-
Fig. 11. Chlorophyll a vs. phosphorus in different en-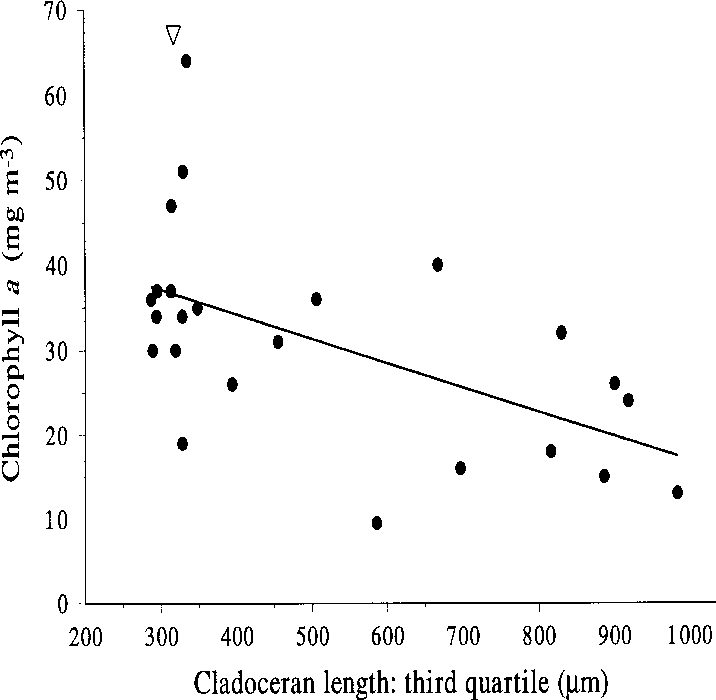 feeding preferences of the crustacean groups.
feeding preferences of the crustacean groups.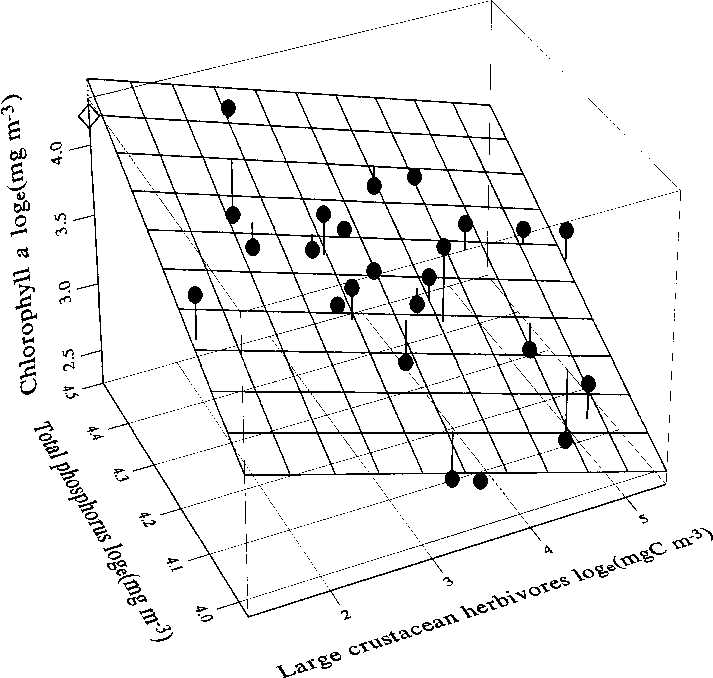 Restoration of the eutrophicated Köyliönjärvi
Fig. 13. Chlorophyll a vs.
Restoration of the eutrophicated Köyliönjärvi
Fig. 13. Chlorophyll a vs.