Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Untitled-4
EFEITO DE BAP E 2,4 – D NA FORMAÇÃO DE CALOS EM DIFERENTES
A. L. OLIVEIRA1 , L. M. HOULLOU-KIDO1, E. A. KIDO1 e A. M. BENKO-ISEPPON1
Resumo - O feijão-caupi (Vigna unguiculata), por ser uma planta de valor significativo para a cultura desubsistência no Brasil, foi utilizado no presente trabalho com o objetivo de verificar o efeito dos hormôniosBAP e 2,4 – D na formação de calos com potencial regenerativo para a cultura in vitro. As sementes defeijão-caupi cv. IPA207 foram utilizadas como fonte dos explantes (eixo embrionário e cotilédones) queforam isolados e colocados em meio MS com diferentes suplementações hormonais. Durante o cultivopôde-se observar diferentes respostas aos hormônios utilizados. No meio I (sem hormônio) houvepredomínio da regeneração dos eixos embrionários (formação de ápices caulinares e raízes), no meio II(2,4 – D) e III (2,4 – D e BAP) observou-se formação de calos nos eixos embrionários. Em relação aoscotilédones, não foi observada uma resposta representativa da formação de calos nos meios II e III. Jáno meio IV (BAP) não houve formação de calos em nenhum dos explantes testados. Estes resultadosindicam que melhor explante para indução de calos em feijão-caupi são os eixos embrionários. Osresultados indicam também que a associação dos fitorreguladores BAP e 2,4 – D favoreceram a formaçãode calos com características organogênicas.
Palavras-chave: reguladores de crescimento, células responsivas, cultura de tecidos de plantas.
BAP AND 2,4 – D EFFECT IN A CALLUS FORMATION IN DIFFERENTS
Abstract - Cowpea (Vigna unguiculata), that have a significant value in Brazilian subsistence agriculture,was used with the aim to study BAP and 2,4-D effect in regenerative callus induction. The cowpeaseeds (cv. IPA-207) were used as explants source (embrionary axis and cotyledons). These explantswere inoculated in MS mediums, with different hormonal supplementation. Different hormonal responseswere observed during culture. In medium I (hormone free) was observed axis embrionary regeneration(shoot tips and root formation), in medium II (2,4 – D) and III (2,4 – D and BAP) was observed callusformation in embrionary axis. Cotyledon callus formation was not significant in medium II and III. Callusformation was not observed in any explants tested in medium IV (BAP). These results suggests thatembrionary axis is the best explants for callus induction in cowpea. These result also indicates afavorable effect of BAP and 2,4-D association on cowpea beans organogenic callus induction.
Keywords: growth regulator, responsive cells, plant tissue culture.
1 Universidade Federal de Pernambuco, CEP: 50670-901, Recife, PE. E-mail:alimadeoliveira@yahoo.com.br; laureenhk@yahoo.com; ederson.kido@gmail.com; celisep@hotlink.com.br
O feijão-caupi [Vigna unguiculata (L.) Walp.] é uma importante leguminosa cultivada nas regiões
Norte e Nordeste do Brasil. Sua importância deriva do seu alto valor nutritivo (proteínas, aminoácidosessenciais, carboidratos, vitaminas, minerais, fibras dietéticas e baixo teor de gordura), além de suacapacidade de se adaptar bem em todos os tipos de solos. O feijão-caupi atua como base alimentar parapopulações de baixa renda do nordeste brasileiro, devido a suas características apropriadas ao plantio,tais como ciclo curto, baixa exigência hídrica e rusticidade para se desenvolver em solos de baixafertilidade (Embrapa, 2003). Além do uso para consumo humano, o feijão-caupi também é utilizado naalimentação animal, adubação verde e proteção do solo. A cultura in vitro de tecidos vegetais é uma dasáreas da biotecnologia que mais contribuiu, nos últimos anos, para o desenvolvimento de novas cultivarescom importantes características agronômicas (resistência a pragas e doenças, precocidade, modificaçãodo porte da planta, etc). A capacidade de indução da desdiferenciação celular é uma das mais importantescaracterísticas exploradas no cultivo in vitro das plantas e encontra-se associada às principais técnicasde melhoramento não convencional (Maluszinsky, 2001; Fehér et al. 2002).Dentre as técnicas maisutilizadas, a seleção in vitro de mutantes induzidos ou provenientes da variação somaclonal e a produçãode transgênicos destacam-se no desenvolvimento de novas cultivares. No entanto, para a utilizaçãodestas metodologias, faz-se necessário o prévio estabelecimento das condições necessárias para aregeneração de plantas via calos (principal tipo de explante utilizado para obtenção de variação somaclonale de transgênicos). Nesse experimento foram utilizadas sementes de feijão-caupi cv IPA207, com oobjetivo de analisar o efeito dos hormônios BAP (6-Benzilaminopurina) e 2,4–D (Ácido 2,4-Diclorofenoxiacético) na formação de calos, com potencial regenerativo, a partir de dois tipos de explantesvegetais (eixos embrionários e cotilédones).
Os experimentos foram realizados no Laboratório de Genética Molecular do Departamento de
Genética da UFPE. Foram utilizadas sementes de feijão-caupi cv IPA207 fornecidas pelo IPA. Inicialmente,neste experimento, foi adotada a seguinte metodologia de desinfestação: três lavagens em água destiladapor 5 minutos cada; imersão em solução de álcool 70% por 1 min, seguida pela imersão em solução dehipoclorito de sódio (NaOCl) a 10% por 20 minutos, seguida de três lavagens com água esterilizada,sendo que, na terceira água, foi adicionada uma cápsula de antibiótico (Keflex). As sementespermaneceram no antibiótico por aproximadamente 18 horas, sendo que, após esse período, os cotilédonese eixos embrionários foram isolados, sob condições assépticas e inoculados em 40 frascos contendo25 ml de meio de cultura para regeneração de feijão-caupi [sais e vitaminas de MS acrescido de sacarose(30 g.L-1) e Agar (6,0 g.L-1)] . Em cada frasco, havia dois cotilédones e um segmento de eixo embrionário. Esse experimento foi montado com quatro tratamentos contendo 10 frascos cada: tratamento I – meiocontrole sem adição de hormônio; tratamento II – adição de 2,4 – D 1 mg.L-1; tratamento III – adição de2,4 – D 1 mg.L-1 e de BAP 0,2 mg. L-1; tratamento IV – adição de BAP 0,2 mg. L-1. Os frascos forammantidos em sala de crescimento com fotoperíodo de 16 h e temperatura controlada (±26o C). Duranteo cultivo foram avaliados o desenvolvimento e o aspecto dos calos formados a partir dos diferentesexplantes e meios de cultura.
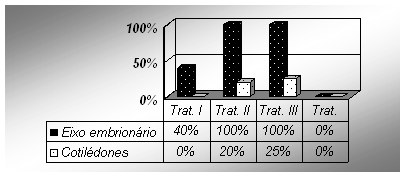
Foram observados os efeitos do BAP e 2,4 – D tanto isolados quanto associados. No tratamento
I (meio de cultura sem hormônios), verificou-se que apenas 40% dos eixos embrionários apresentaramsinais de formação de pequenos calos em uma das extremidades do explante. Nos eixos embrionários
restantes (60%) foi observada regeneração direta (sem a formação de calo) de ápices caulinares e/ouraízes (Figuras 1 e 2). Nesse mesmo tratamento, nenhum dos cotilédones apresentou sinais de regeneraçãoou de formação de calos. Já no tratamento II, foi observada a formação de calos em 100% dos eixosembrionários. No entanto, 20% dos explantes formaram pequenos calos em uma das extremidades,enquanto que nos 80% restante houve formação de calo em todo explante (Figuras 1 e 3). Matsumoto& Yamaguchi (1991) sugerem que esse tipo de resultado pode ser causado pela existência deheterogeneidade fisiológica dos explantes durante o cultivo in vitro. Estes resultados também indicamque os eixos embrionários apresentam o melhor nível de resposta ao 2,4 – D para a formação de caloscom potencial organogênico. A diferença na resposta ao fitorregulador provavelmente está correlacionadaa diferenças na quantidade de células responsivas existentes entre os dois tipos de explantes testados(eixos embrionários e cotilédones). Segundo dados da literatura, um dos melhores tecidos para a formaçãode calos é a epiderme de embriões.No entanto, o desenvolvimento de células e calos embriogênicos épositivamente influenciado pela composição do meio de cultura (Meijer et al., 1999).
Figura 1. Formação de calos sob efeito de diferentes tratamentos.
Figura 2. Regeneração dos tecidos vegetais no tratamento I. Figura 3. Formação de calo em todo o eixo embrionário.
Apenas 20% dos cotilédones apresentaram algum sinal da formação de calos na extremidade em
que o embrião estava ligado. Esse resultado pode ser devido à presença de restos da epiderme doembrião no cotilédone. No tratamento III, o qual apresenta uma combinação entre BAP e 2,4 – D,verificou-se a formação de calos em 100% dos eixos embrionários. Nesse tratamento foi observada aconversão de todo o eixo embrionário em calo, não havendo a restrição deste a apenas uma região doexplante. Com relação aos cotilédones, a resposta foi de 25% para início da formação de calos(Figuras 1 e 4). Infere-se então que a associação entre esses dois hormônios age de forma positiva
tanto no nível de respostas de eixo embrionário quanto de cotilédones. No tratamento IV (meio decultura suplementado apenas com BAP), nenhum dos explantes iniciou a formação de calos. Houveapenas a regeneração direta de ápices caulinares e/ou de raízes em 10% dos eixos embrionários. Nessetratamento, foi observado também que 70% dos eixos embrionários apresentaram intumescimento dostecidos (Figura 1 e 5) sem a ocorrência de regeneração ou formação de calo. Este resultado indica queo BAP sozinho não consegue induzir a formação de calos.
Figura 4. Início da formação de calo em cotilédone. Figura 5. Eixo embrionário intumescido.
O 2,4 - D sozinho ou em associação com o BAP tem a capacidade de induzir a formação de calos
O eixo embrionário apresenta-se como o melhor tipo de explante a ser utilizado por ser mais
O BAP sozinho não cumpre o papel de indutor de formação de calos, porém apresenta resposta
positiva no que diz respeito à regeneração dos explantes.
Ao IPA – Empresa Pernambucana de Pesquisa Agropecuária – por fornecer a cv IPA207 e assim
facilitar o nosso trabalho, ao Professor Dr. Éderson A. Kido responsável pelo laboratório de GenéticaMolecular do Departamento de Genética da Universidade Federal de Pernambuco, e à Dra. LaureenMichelle H. Kido pelas orientações.
Embrapa Meio-Norte: Sistemas de Produção, 2 ISSN 1678-8818 Versão Eletrônica Jan/2003. Disponí-vel em: http://sistemasdeproducao.cnptia.embrapa.br/FontesHTML/Feijao/FeijaoCaupi/importancia.htm Acesso em: 20 mar. 2006.
FEHÉR, A.; PASTERNAK, T.; DUDITS, D. Activation of embryogenic cell division in leaf protoplast-derived alfalfa cells: the role of auxin and stress. Acta Biologica Szegediensis, v. 46, p.13-14, 2002. http://www.sci.u-szeged.hu/ABS.
MATSUMOTO, K.; YAMAGUCHI, H. Induction and selection of aluminium tolerance in the banana. Viena: IAEA, 1991. p. 249-255.
MEIJER, E. A.; DE VRIES, S. C.; MORDHORST, A. P. Co-culture with Daucus carota somatic embryosreveals high 2,4-D uptake and release rates of Arabidopsis thaliana cultured cells. Plant Cell Reports, v. 18, p.656–663, 1999.
MURASHIGE T.; SKOOG F. A revised medium for rapid growth and bioassays with tobacco tissueculture. Physiologia Plantarum, v. 15, n. 4, p. 473-497, 1962.
Sex Differences in Jealousy: An Evolutionary Perspective on Online Infidelity R E. G1This study examined whether sex differences in jealousy would generalize to onlineinfidelity. Based on the evolutionary psychological explanation for sex differences injealousy (ancestral men’s challenge of paternal uncertainty vs. ancestral women’schallenge of ens
New aspects of the renin–angiotensin system: angiotensin-converting enzyme 2 – a potential target for treatment ofhypertension and diabetic nephropathyDaniel BaMarı´a Jose´ and Jan WaDivision of Nephrology and Hypertension, Departmentof Medicine, The Feinberg School of Medicine,Whereas angiotensin-converting enzyme promotes the formation of angiotensin II,Northwestern University, Chicag


 restantes (60%) foi observada regeneração direta (sem a formação de calo) de ápices caulinares e/ouraízes (Figuras 1 e 2). Nesse mesmo tratamento, nenhum dos cotilédones apresentou sinais de regeneraçãoou de formação de calos. Já no tratamento II, foi observada a formação de calos em 100% dos eixosembrionários. No entanto, 20% dos explantes formaram pequenos calos em uma das extremidades,enquanto que nos 80% restante houve formação de calo em todo explante (Figuras 1 e 3). Matsumoto& Yamaguchi (1991) sugerem que esse tipo de resultado pode ser causado pela existência deheterogeneidade fisiológica dos explantes durante o cultivo in vitro. Estes resultados também indicamque os eixos embrionários apresentam o melhor nível de resposta ao 2,4 – D para a formação de caloscom potencial organogênico. A diferença na resposta ao fitorregulador provavelmente está correlacionadaa diferenças na quantidade de células responsivas existentes entre os dois tipos de explantes testados(eixos embrionários e cotilédones). Segundo dados da literatura, um dos melhores tecidos para a formaçãode calos é a epiderme de embriões.No entanto, o desenvolvimento de células e calos embriogênicos épositivamente influenciado pela composição do meio de cultura (Meijer et al., 1999).
restantes (60%) foi observada regeneração direta (sem a formação de calo) de ápices caulinares e/ouraízes (Figuras 1 e 2). Nesse mesmo tratamento, nenhum dos cotilédones apresentou sinais de regeneraçãoou de formação de calos. Já no tratamento II, foi observada a formação de calos em 100% dos eixosembrionários. No entanto, 20% dos explantes formaram pequenos calos em uma das extremidades,enquanto que nos 80% restante houve formação de calo em todo explante (Figuras 1 e 3). Matsumoto& Yamaguchi (1991) sugerem que esse tipo de resultado pode ser causado pela existência deheterogeneidade fisiológica dos explantes durante o cultivo in vitro. Estes resultados também indicamque os eixos embrionários apresentam o melhor nível de resposta ao 2,4 – D para a formação de caloscom potencial organogênico. A diferença na resposta ao fitorregulador provavelmente está correlacionadaa diferenças na quantidade de células responsivas existentes entre os dois tipos de explantes testados(eixos embrionários e cotilédones). Segundo dados da literatura, um dos melhores tecidos para a formaçãode calos é a epiderme de embriões.No entanto, o desenvolvimento de células e calos embriogênicos épositivamente influenciado pela composição do meio de cultura (Meijer et al., 1999).
 tanto no nível de respostas de eixo embrionário quanto de cotilédones. No tratamento IV (meio decultura suplementado apenas com BAP), nenhum dos explantes iniciou a formação de calos. Houveapenas a regeneração direta de ápices caulinares e/ou de raízes em 10% dos eixos embrionários. Nessetratamento, foi observado também que 70% dos eixos embrionários apresentaram intumescimento dostecidos (Figura 1 e 5) sem a ocorrência de regeneração ou formação de calo. Este resultado indica queo BAP sozinho não consegue induzir a formação de calos.
tanto no nível de respostas de eixo embrionário quanto de cotilédones. No tratamento IV (meio decultura suplementado apenas com BAP), nenhum dos explantes iniciou a formação de calos. Houveapenas a regeneração direta de ápices caulinares e/ou de raízes em 10% dos eixos embrionários. Nessetratamento, foi observado também que 70% dos eixos embrionários apresentaram intumescimento dostecidos (Figura 1 e 5) sem a ocorrência de regeneração ou formação de calo. Este resultado indica queo BAP sozinho não consegue induzir a formação de calos.