Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Pii: s1522-6514(01)80017-3
International Journal of Phytoremediation: Vol. 3, No. 2, pp. 173–187 (2001)
The Effect of Recombinant Heavy Metal-Resistant Endophytic Bacteria on Heavy Metal Uptake by Their Host Plant C. Lodewyckx,1,2 S. Taghavi,1 M. Mergeay,1 J. Vangronsveld,2 H. Clijsters,2 and D. van der Lelie1* 1Vito, Environmental Technology, Boeretang 200, B-2400 Mol, Belgium; 2Department SBG, Environmental Biology, Limburgs Universitair Centrum, Universitaire Campus, B-3590 Diepenbeek, Belgium
Corresponding author: D. van der Lelie, Tel.: 0032 14 33 51 66. Fax.: 0032 14 580523. email: Niels.vanderlelie@vito.be
ABSTRACT
The ncc-nre nickel resistance system of Ralstoniametallidurans 31A was effi-
ciently expressed in Burkholderia cepacia L.S.2.4 and Herbaspirillum seropedicaeLMG2284. The heterologous expression of ncc-nre encoded nickel resistance wasaccompanied by nickel removal from the culture medium. B. cepacia L.S.2.4::ncc-nre and H. seropedicae LMG2284::ncc-nre were able to remove 35 and 15% nickel,respectively. The capacity to remove nickel through sequestration or bio-precipita-tion processes and consequently lowering the free nickel concentration could offerinteresting benefits for these endophytic bacteria and their host plants. Once inocu-lated in their host plant, they could possibly alter the nickel speciation and thereforedecrease the free ions and thus toxic concentration for the plant metabolism. Lupinusluteus L, when grown on a nickel enriched substrate and inoculated with B. cepaciaL.S.2.4::ncc-nre, showed a significant increase (30%) of nickel concentration in theroots, whereas the nickel concentration in the shoots remained comparable with thatof the control plants. The inoculation of Loliumperenne (cv Atlas) with the nickelresistance derivative of H. seropedicae LMG2284::ncc-nre resulted in a significantdecrease of the nickel concentration in the roots (11%) as well as in the shoots (14%). As this phenomenon was also observed in the Loliumperenne plants inoculated withthe wild-type strain LMG2284, the nickel resistance characteristics probably are notresponsible for the altered nickel uptake observed. KEY WORDS: phytoextraction, Burkholderiacepacia, Herbaspirillumseropedicae, Lupinusluteus, Loliumperenne, heterologous expression.
1522-6514/01/$.50 2001 by CRC Press LLC
Lodewyckx et al. I. INTRODUCTION
Living plants have the ability to easily assimilate heavy metals that are essential
in trace concentrations for their growth and development and this has been used todevelop phytoextraction as a remediation technique (Baker and Walker, 1990;Mehra and Farago, 1994). Although offering some interesting benefits comparedwith the traditional remediation techniques, phytoextraction still has its limitations. A suitable plant used in extraction of heavy metals should possess several charac-teristics, which are rarely found within one plant species. For this reason, differentstrategies are currently being investigated in order to improve crops forphytoextraction processes (Cunningham and Berti, 1993; Cunningham and Ow,1996; Burd et al., 1998; Arazi etal., 1999; Brewer et al., 1999). In this context weinvestigated the possible interaction between plant associated endophytic bacteriaand their host plant with respect to the uptake and accumulation of heavy metals. Endophytic bacteria are defined as those microorganisms that are able to enterplant tissues and to establish themselves inter- and intracellularly (Di Fiori and DelGallo, 1995). They have the ability to establish an active relationship with theirhosts and can be defined as colonists (Kloepper and Beauchamp, 1992; Kloepperet al., 1992). The introduction and heterologous expression of known heavy metalresistance genes in these endophytic bacteria might have an effect on the uptakecapacities of heavy metals by their host plant. In fact, Salt et al. (1999) showed thatCd-tolerant rhizobacteria were able to promote Cd precipitation processes near theroot surface of Indian mustards plants and consequently decreased the toxic effectsof the metal cation for the roots. Previous studies have shown that several mecha-nisms can be responsible for bacterial heavy metal resistance, for example, block-ing the entry of toxic ions in the cells, intracellular sequestration of the metals bymetal binding proteins, enzymatic conversion of the metal to a less toxic form, andenergy-driven efflux systems for cations and anions encoded by resistance genes,such as the czc, cnr, ncc, cad, and ars operons (Mergeay, 1997; Taghavi et al.,1997). Bioprecipitation and sequestration processes also seem to take place whenbacteria are equipped with efflux mechanisms. This phenomenon was observed incultures of Ralstoniametallidurans CH34 (previous Alcaligeneseutrophus CH34)when grown in the presence of high concentrations of Cd or Zn and attributed tothe action of the czc resistance operon on the pMOL30 plasmid (Diels et al., 1995). Such bioprecipitation and sequestration characteristics could offer interesting ben-efits for the bacteria, and in the case of endophytic bacteria the speciation of theheavy metals might be altered in the host plant from a free to a less available form. This could lead to a reduced toxicity of the heavy metals on plant metabolism. Inorder to test this hypothesis, Burkholderiacepacia and Herbaspirillumseropedicaewere selected as endophytic bacteria. Some B. cepacia strains have been reportedas facultative endophytes of lupine plants (K. Ophel Keller, personal communica-tion) or were found to colonize roots of various maize cultivars (Hebbar etal.,1992a; Hebbar etal., 1992b). H. seropedicae strains are obligate endophytes ofmany genera of the Gramineae and do not seem to survive in uncropped soils(Baldani etal., 1992, 1996; Olivares etal., 1996). Both species were used toinvestigate the heterologous expression of heavy metal efflux systems, as well as
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake
the effects of this expression on heavy metal accumulation and translocation byplants after inoculation with the recombinant endophytic strains. II. MATERIALS AND METHODS A. Bacterial Strains, Plasmids, and Media
The bacterial strains and plasmids used in this study are listed in Table 1. B. cepacia L.S.2.4 was originally isolated from surface sterilized stems of lupinegrowing on a wheat-pasture rotation soil from Kapunda, South Australia. The endo-phytic character of this strain was confirmed by growing Lupinusluteus L. seedlingsin the presence of 108 CFU/ml of the nickel resistant variant (L.S.2.4::ncc-nre) addedto the perlite growth medium. Surface sterilized and macerated root and shoot tissueof 14-day-old plants harbored 1.4 × 106 CFU/gFW and 4.3 × 103 CFU/gFW, respec-tively, when plated on selective media containing Ni. H. seropedicae LMG2284 was obtained from the Laboratory of Microbiology in
Gent, Belgium. Loliumperenne cv Atlas, selected to check the ability of this strainto establish itself endophytically, also proved to function as a host plant. After growthunder similar conditions as mentioned for Lupinusluteus L., 6.8 × 106 CFU/gFW and1.1 × 105 CFU/gFW of the nickel-resistant variants (LMG2284::ncc-nre) wererecovered from surface-sterilized and macerated root and shoot tissue, respectively,when plated on nickel containing selective media.
All strains were grown at 30°C in a 284 minimal medium (Mergeay et al., 1985)
supplemented with 0.2% gluconate as a carbon source. Escherichia coli strains weregrown at 37°C in Luria Broth (LB) medium (Sambrook etal., 1989). Resistance toheavy metal salts was tested in Tris-minimal medium (Mergeay et al., 1985). Mini-mum Inhibitory Concentrations were determined by growing the strains in thesemedia supplied with increasing concentrations (0.1–5 mM) of the metal to be tested. At least four replications were done for each strain. B. Heterologous Expression of Cu, Cd, Co, Ni, Pb, and Zn Resistance
For the introduction of plasmids in B. cepacia L.S.2.4 and H. seropedicae
LMG2284, triparental matings were performed. E. coli CM404 (pRK2013) (Figurskyetal., 1982) was used as a helper strain for plasmid mobilization. The E. coli strainsCM1366 (czc), CM1623 (czr), CM1666 (pbr), CM1684 (cop) and CM1962 (ncc-nre)were used as donors. LMG2284 transconjugants were selected on 284 minimalmedium supplemented with 20 µg/ml tetracycline, while L.S.2.4 transconjugantswere selected on 284 minimal medium supplemented with 150 µg/ml tetracycline. For matings with CM1962, 100 µg/l kanamycine was used for selection.
Due to the instability of the introduced plasmids in B. cepacia and H. seropedicae,
miniTn5-Km1-based transposons (de Lorenzo et al., 1990) containing ncc and nrewere constructed in order to have the expressed nickel-resistant determinants insertedin the chromosome stabily (Taghavi etal., 2001 ). This resulted in plasmid pMOL1554,which was used as donor for the mobilization of the pUTminiTn5-Km1::ncc-nre toB. cepacia and H. seropedicae. Transconjugants were selected on 284 minimalmedium containing either Km or Ni. Lodewyckx et al. TABLE 1. Bacterial strains and plasmids used. Strain or plasmid Relevant characteristics Source/ Ref.
Wild type, pMOL28, pMOL30 Mergeay et al., 1985
Wild type, endophyte of lupine K. Ophel-Keller
(pKT240-ncc-nre) Apr, Kmr, Nir, Dong etal., 1998
(pLAFR3-czc) Tcr, Cdr, Znr, Cor, van der Lelie etal.,
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake C. Nickel Removal
Strains B. cepacia L.S.2.4::ncc-nre, H. seropedicae LMG2284::ncc-nre, and
R. metallidurans 31A were grown at 30°C for 7 days in SVR medium (294 mg Na-
β-glycerol-phosphate, 1 g NH NO , 0.2 g MgSO .7H O, 0.01 g FeSO .7H O, 0.1g
CaCl .2H O, 20 g C H O per liter) supplemented with 0.4 mM NiCl until the
stationary growth phase. Absorbency at 660 nm (A
samples were taken from the culture, centrifuged (6000 rpm for 10'; Sorvall RC5C)and the supernatant filtered through a 0.45-µm filter unit (Gelman sciences). Subse-quently, the Ni concentrations were determined using Atomic Absorption Spectros-copy (A.A.S.). D. Plant Inoculation Experiments 1. Surface Sterilization of Seeds Lupinus luteus L. seeds were surface sterilized by shaking for 30 min in a
commercial NaOCl solution supplemented with 1% active chloride (Dumortier,Merksem) and one droplet (per 100 ml) of Triton X-100®, rinsed thoroughly in sterilewater for 30 min, and incubated during 3 days at 30°C on LB medium in order to teststerility.
The same procedure was followed for the surface sterilization of the Loliumperenne seeds, except that the NaOCl solution contained 3% active chloride, and theseeds were shaken for 60 min. 2. Preparation of Bacterial Inocula
Overnight cultures of L.S.2.4::ncc-nre and LMG2284::ncc-nre were grown in LB
medium supplemented with 4 mM NiCl at 30°C on a rotary shaker, centrifuged at 6000
rpm during 15' (Sorvall, RC5C), washed twice in a 0.85% NaCl solution, diluted andplated on LB medium with and without 4 mM NiCl in order to test the purity of the
solution and the presence of the nickel resistance. Wild-type strains L.S.2.4 andLMG2284 were grown under the same conditions without addition of NiCl . 3. Inoculation and Plant Growth Conditions
Surface-sterilized Lupinusluteus L. seeds were grown in sterile plastic jars (800
ml) (Duchefa, Labconsult) completely filled with perlite and saturated with 1/2sterile Hoagland’s solution. Perlite was chosen as plant growth substrate because itcan be easily sterilized and provides the roots with moisture, nutrients, and goodaeration due to the large surface area and the physical shape of each particle. Thebacterial inoculum was added at a concentration of 108 Colony Forming Units (CFU)per ml (in NaCl 0.85%), whereas for the noninoculated control plants the sameamount of NaCl 0.85% was added. Five seeds were planted in each jar and thancovered with tinfoil in order to prevent contamination and dispersion of the inocu-lated bacteria through the air. Once the germinated seeds were able to grow throughthe holes that were made in the tinfoil, they were grown with a 12-h light and darkcycle, 65% relative humidity, and at a constant temperature of 22°C. In order to keepthe perlite moistened sterile water was added every 4 days. Lodewyckx et al.
The surface sterilized Loliumperenne seeds were grown under the same condi-
tions as the lupine seeds except that the plastic jars were filled with 200 ml perlite. Seeds were randomly spread over the available area in order to guarantee a similarbiomass production as for the lupine seedlings. 4. Harvest of Plant Material and Nickel Determination
After 21 days, the Lupinusluteus and Loliumperenne plants were harvested. For 30
plants, the length of roots and shoots was measured separately as well as the producedbiomass. Fresh root and shoot material was vigorously washed with distilled water inorder to remove all trace amounts of nickel that could be present on the surface of theplant material. After drying the root and shoot biomass for 24 h at 65°C, it wasmacerated with a mortar and pestle and microwave digested in supra pure HNO (70%)
in order to release all nickel, whose concentration was determined by means of A.A.S. E. Statistical Analysis
The morphological data as well as the nickel concentrations in roots and shoots
of Lupinusluteus and Loliumperenne were analyzed by a one-way analysis ofvariance (ANOVA). The different treatments were compared by means of a Tukeyhonest significant difference (HSD) test. Significance was tested at the level ofP = 0.05. III. RESULTS A. Heterologous Expression of Cu, Cd, Co, Ni, Pb, and Zn Resistance
The cop (copper resistance), pbr (lead resistance), ncc-nre (nickel-cadmium-cobalt resis-
tance), and czr (cadmium-zinc resistance) heavy metal resistance operons cloned in broad hostrange IncQ (pKT240) or IncP (pLAFR3) plasmids were transferred into B. cepacia L.S.2.4and H. seropedicae LMG2284 by triparental matings. After selection for antibiotic resistance,heterologous expression of heavy metal resistance was examined. The best expressed markerfor both B. cepacia and H. seropedicae was the nickel resistance of ncc-nre, whose expressionresulted in an increase of the MIC value for Ni from 0.4 or 0.6 mM to 2 mM for H. seropedicaeLMG2284 and B.cepacia L.S.2.4, respectively. Low expression of cop-encoded Cu resistancewas seen in H. seropedicae, while no heterologous expression of the pbr and czr operons wasobserved. Because the ncc-nre nickel resistance was the best marker in both strains, wecontinued to study its heterologous expression in more detail. Due to the instability of thepMOL222 plasmid, a miniTn5 transposon containing ncc-nre was used to obtain a stableinsertion of the nickel resistance determinant into the chromosome. For both B. cepacia andH. seropedicae, the minitransposon carrying ncc-nre proved to be stable when inserted in thechromosome (no loss of nickel resistance after growth for 100 generations under nonselectiveconditions) and gave a MIC for Ni of 2 mM on 284 minimal medium in both strains. B. Nickel Removal
The nickel-resistant endophytic strains L.S.2.4::ncc-nre and LMG2284::ncc-nre
were tested for their ability to remove nickel from the growth medium. As a positive
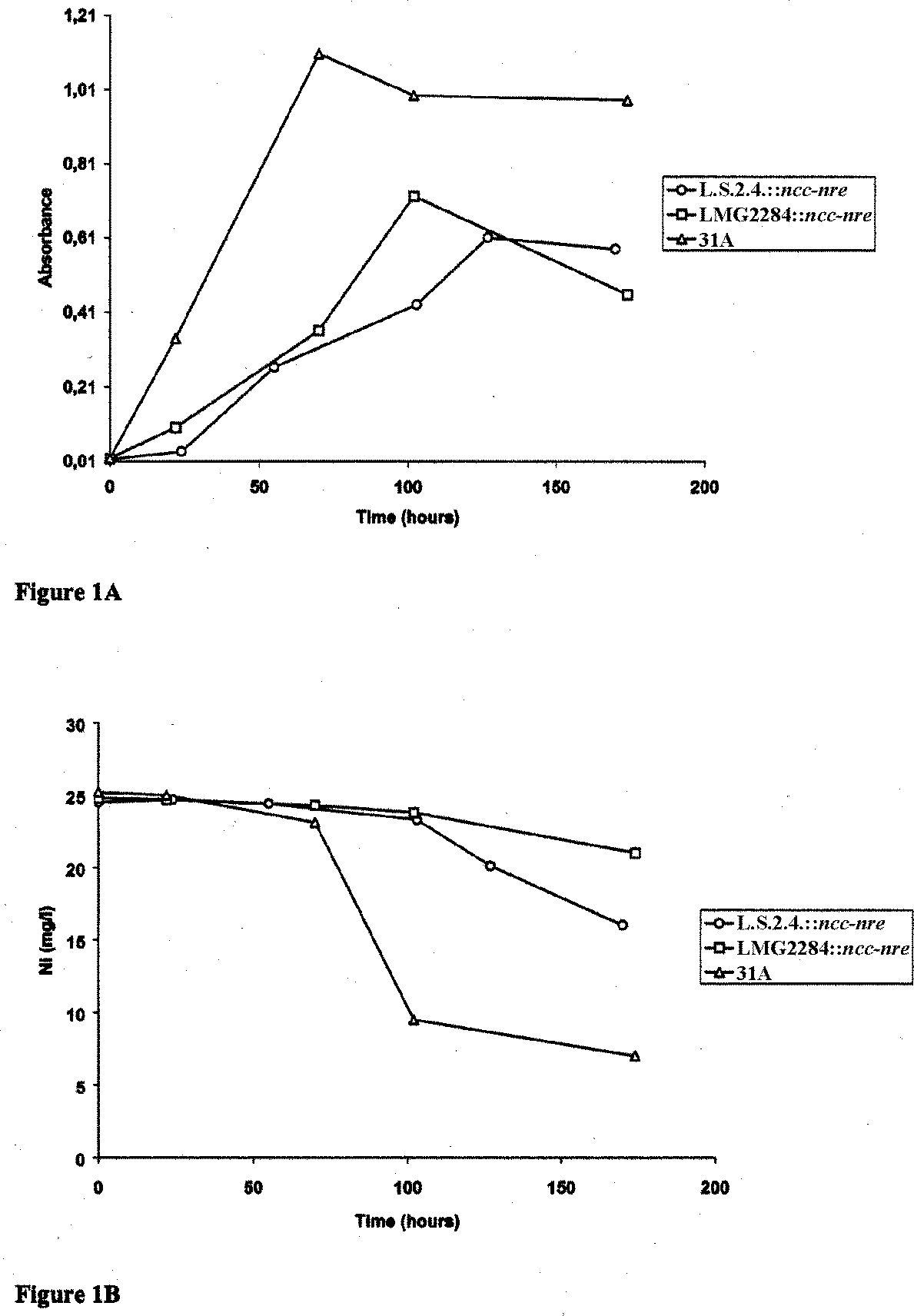
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake Figure 1(A). Optical Density (OD
) in function of time for R. metallidurans 31A, B. cepacia
L.S.2.4.::ncc-nre, and H. seropediacae LMG2284::ncc-nre cultured in SVRmedium supplemented with 0.4 mM NiCl . The data are representative for three
separate and independent experiments. (B). Concentration of Ni (mg/l) in the culture supernatant over time for R. metallidurans 31A, B. cepacia L.S.2.4.::ncc- nre, and H. seropediacae LMG2284::ncc-nre, grown in SVR medium supple- mented with 0.4 mM NiCl . The data are representative for three separate and Lodewyckx et al.
control, the wild-type strain R. metallidurans 31A was used, which contains copiesof both ncc and nre on its endogenous megaplasmids pTOM8 and pTOM9. Growingthe strains for 180 h in minimal SVR medium supplemented with 0.4 mM NiCl2revealed that the R. metallidurans 31A strain was able to grow at a higher rate thanthe endophytic strains (Figure 1A). This was probably due to the multiple copies ofthe nickel resistance determinants and perhaps other important functions of nickel-resistance on pTOM8 and pTOM9. These features are also important when the nickelremoval capacity is considered (Figure 1B). R. metallidurans 31A strain removed74% of the nickel present in the growth medium, while B. cepacia L.S.2.4::ncc-nreand H. seropedicae LMG2284::ncc-nre only removed 35 and 15% of nickel, respec-tively. The pH of the growth medium also increased over time. The pH of theR. metallidurans 31A culture was increased from pH 7 to pH 9, while the pH of theL.S.2.4::ncc-nre and LMG2284::ncc-nre changed from 7 to 8 (data not shown). C. Nickel Toxicity for Lupinus luteus L. and Lolium perenne (cv. Atlas)
Preliminary experiments were performed to test the sensitivity of Lupinus luteus L.
seedlings for nickel in order to decide which concentration would be suitable for theinoculation experiments. From these experiments it was decided to test the effect of0.25 mM NiCl because this concentration had a visible but nonlethal effect on the
growth response of the Lupinus seedlings. Root and shoot biomass and length weresignificantly reduced when nickel was added to the perlite (data not shown).
As was observed for the LupinusluteusL. seedlings, the growth of Loliumperenne seedlings was also reduced when treated with increasing NiCl concentra-
tions. Root and shoot biomass and length significantly decreased with increasingnickel concentrations added to the growth substrate (data not shown). In order tocompare the effects in both plant species, we decided to use the same concentration(0.25 mM NiCl ) as for the experiments with the lupine plants. D. Inoculation of Lupinus luteus L. with B. cepacia L.S.2.4 and Its Nickel- Resistant Derivative L.S.2.4::ncc-nre
Wild-type strain L.S.2.4 and its nickel-resistant derivative L.S.2.4::ncc-nre were
inoculated in perlite and the Lupinus seedlings were grown on this substrate for 21days under controlled environmental conditions. Noninoculated sterile plants wereused as controls. In the absence of NiCl , no difference in growth response was
observed between the 21-day-old noninoculated control plants and the inoculatedlupine plants when the shoot biomass and length were considered (Table 2). Theroots seemed to be slightly but significantly affected in their growth when B. cepaciawas added as a wild-type strain or as the nickel-resistant derivative, indicating thatthe presence of B. cepacia L.S.2.4 has a minor effect on the root development in theabsence of nickel (Table 2). The nickel concentration in both roots and shoots wasmeasured with A.A.S but was below the detection limit (<2.5 mg/kg DW).
The addition of 0.25 mM NiCl to the perlite resulted in a decrease of the growth
parameters when compared with the treatment without NiCl , suggesting a toxic
effect of the nickel cations. Table 2 shows the morphological parameters of the lupineplants with the different inocula in the presence of 0.25 mM NiCl . The presence of
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake TABLE 2: Morphological parameters of Lupinus luteus l. and Lolium perenne Plants for the Different Inocula and the Different Nickel Treatments Root biomass Root length Shoot biomass Shoot length Lupinus luteus L. Lolium perenne
L.S.2.4 and LMG2284 were inoculated as wild-type strains, whereas L.S.2.4::ncc-nreand LMG2284::ncc-nre were inoculated as its nickel-resistant derivatives in Lupinusand Loliumperenne. respectively. Data are mean values of 30 replicate seedlings ±S.D. Different superscripted letters indicate values that are statistically different(P < 0.05). Lodewyckx et al. B. cepacia, both the wild-type or its nickel-resistant derivative, did not influence thegrowth of the plants. No significant differences were observed when root and shootbiomass and length were measured. However, a different response in nickel accumu-lation was observed when the nickel concentration in the roots was compared for thedifferent treatments (Figure 2A). A significantly higher total nickel concentrationwas measured in the lupine roots inoculated with the nickel-resistant B. cepaciaL.S.2.4::ncc-nre, while the noninoculated control plants and the plants inoculatedwith the wild-type strain L.S.2.4 had similar but lower nickel contents. In contrast tothe roots, the nickel concentration in the shoots was comparable for the differenttreatments (Figure 2A). E. Inoculation of Lolium perenne with H. seropedicae LMG2284 and Its Nickel-Resistant Derivative LMG2284::ncc-nre
In the absence of nickel, the 21-day-old Loliumperenne plants did not show
significant differences in their shoot growth parameters when the different treatmentswere compared (Table 2). The presence of H.seropedicae LMG2284 did not influ-ence the growth conditions of the plant except for the root development (Table 2). Slight but significantly lower root lengths were measured when the plants wereinoculated with the wild-type and the nickel-resistant derivatives of LMG2284. Thenickel concentration in both roots and shoots was below the detection limit of 2.5 mg/kg DW.
The Loliumperenne plants did suffer from the 0.25 mM NiCl treatment. Root and
shoot biomass and length strongly decreased compared to the untreated control(Table 2). As observed for the 0 mM NiCl treatment, the different inocula actually
did not seem to have major effects on plant growth (Table 2). In contrast, analysis of the nickel content in the roots and shoots revealed that the noninoculated Lolium perenne plants accumulated more nickel when compared with the inoculated plants (Figure 2B). The addition of the wild-type LMG2284 and its nickel-resistant deriva- tive leads to a significantly lower nickel content in the roots when compared with the noninoculated plants, and this was even more pronounced for the plants inoculated with the nickel-resistant LMG284::ncc-nre. The same phenomenon was observed in the shoots except that the nickel content was not significantly different for both wild- type LMG2284 and nickel-resistance derivative LMG2284::ncc-nre inoculation treat- ments. IV. DISCUSSION
The introduction of heavy metal resistance systems from bacteria of polluted
environments in the selected endophytic strains B.cepacia L.S.2.4 and H.seropedicaeLMG2284 was performed in order to test the effect of such bacteria in uptake andtolerance for heavy metals in their host plants. Among the heavy metal resistancesystems used in this study, only the ncc-nre system encoding for Ni, Cd, and Coresistance in R.metallidurans 31A was clearly expressed in the endophytic hosts. Due to the instability of the pMOL222 plasmid, we introduced the ncc-nre genes inthe endophytic strains by means of miniTn5 transposons. This approach resulted in
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake Figure 2 (A). Ni concentration (mg/kg DW) in roots and shoots of Lupinus luteus L. plants
for the different inocula and the 0.25 mM NiCl treatment. L.S.2.4 was inocu-
lated as the wild-type strain and L.S.2.4::ncc-nre as its Ni-resistant derivative. Data are mean values of three replicate samples ± S.D. Different letters indicate values that are statistically significant (P < 0.05). (B). Ni concentration (mg/kg DW) in roots and shoots of Lolium perenne. plants for the different inocula and the 0.25 mM NiCl treatment. LMG2284 was inoculated as the wild-type strain
and LMG2284::ncc-nre as its Ni-resistant derivative. Data are mean values ofthree replicate samples ± S.D. Different letters indicate values that are statisti-cally significant (P < 0.05).*
Lodewyckx et al.
stable nickel-resistant strains that were used in the subsequent nickel-removal andinoculation experiments.
The removal of nickel and alkalization of the growth medium as observed for the
R. metallidurans 31A culture and to a lesser extent for the endophytic strains wassimilar to the removal of zinc and cadmium observed with the czc system ofR. metallidurans CH34 (Diels et al., 1995). In the case of cadmium removal by czc,alkalization of the medium, the formation of metal carbonates from CO produced by
the cell metabolism, and precipitation of the metal carbonates on the cell surface arethought to be the main factors (Diels etal., 1993). The mechanism of nickel removalin our study is unknown, although we demonstrated that R. metallidurans cells areable to sequestrate nickel during the beginning of the stationary growth phase due tothe expression of ncc, which was postulated to act as a cation-proton antiportersystem (Nies, 1992).
Table 2 shows that the measured morphological parameters from 21-day-old
Lupinus and Loliumperenne plants whether inoculated with their respective endo-phytic bacteria were the same except for the root length parameter, where a slightlylower but significant difference was measured between the control plants and theinoculated plants. This suggests that the endophytic strains did not act like PlantGrowth Promoting Bacteria (PGP), having a positive effect on plant growth, or aspathogens, having a negative effect or disease symptoms. This agrees with Bevivinoetal. (1998), who showed that bacterization of Zeamays L. seeds with B. cepaciaisolates had no significant influence on plant development. Herbaspirillum spp. werereported as being good colonists of graminaceous plants, without causing symptomsof plant disease (Reinhold-Hurek and Hurek, 1998).
When the nickel content in the roots and shoots is considered, Loliumperenne and
Lupinus plants responded differently. Figure 2A shows that Lupinus is able toaccumulate higher levels of nickel in its roots, while Loliumperenne seems to be abetter translocator of nickel to the shoots (Figure 2B). The effect of the inoculationon the nickel content in the roots and shoots, when compared with the noninoculatedcontrol plants, differed for the selected B. cepacia and H. seropedicae strains. Thecolonization by the nickel-resistant B. cepacia L.S.2.4::ncc-nre causes a significantincrease in the nickel concentration in the roots. This phenomenon could be attributedto the heterologous expression of the ncc-nre system in the B. cepacia L.S.2.4::ncc-nre strains and the subsequent sequestration processes, which are presumed to occurand that could affect the free and therefore toxic nickel fraction in the roots. Incontrast to the root, the nickel concentration in the shoots does not seem to beinfluenced by the presence of the nickel-resistant B. cepacia L.S.2.4::ncc-nre. Apossible explanation is the preferential colonization of the Lupinus roots. Anotherreason might be that the nickel ions are complexed in the roots, and consequentlytheir transfer to the shoots is blocked.
The presence of nickel-resistant H. seropedicae in Loliumperenne seems to have
the same effect on the nickel concentration in both the roots and shoots. Comparedwith the noninoculated control plants, the nickel content decreased slightly in the 21-day-old Loliumperenne roots and shoots when inoculated with the wild-type as wellas the nickel-resistant derivative of H. seropedicae. This suggests that the occurrenceof these strains, both nickel-sensitive and nickel-resistant, could be responsible for
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake
a kind of protection, which results in a decreased nickel uptake by the host plant. Thefact that only minor nickel complexation was observed in growth cultures ofLMG2284::ncc-nre might imply that a successful inoculation in its host plant wouldnot change nickel uptake and accumulation significantly.
The experiments with inoculated plants reported here suggest that B. cepacia and
H. seropedicae as endophytic wild-type and nickel-resistant bacteria are responsiblefor altered nickel uptake capacities in their respective host plant. However, thispresence does not contribute to an improvement of the nickel translocation to theshoots, which would have been an important feature to improve the efficiency ofphytoextraction processes focussing the aboveground plant tissues. It should also bementioned that these experiments were performed under laboratory conditions and ina sterile environment without any interference of competing microorganisms that arepresent in soil. In situ conditions should certainly be studied as well in order toevaluate the behavior of these endophytic bacteria and their potential contribution tothe heavy metal uptake and translocation capacities of its host plant. REFERENCES Arazi, T., Sunkar, R., Kaplan, B., and Fromm, H. 1999. A tobacco plasma membrane
calmodulin-binding transporter confers Ni2+ tolerance and Pb2+ hypersensitivity in transgenic plants. The Plant Journal20: 171–182.
Baldani, V.L.D., Baldani, J.I., Olivares, F., and Döbereiner, J. 1992. Identifation and ecology
of Herbaspirillum seropedicae and the closely related Pseudomonas rubrisubalbicans. Symbiosis 13: 65–73.
Baldani, J.I., Baldani, V.L.D., Olivares, F.L., Kirchof G., Hartman A., Pot B., Hoste B.,
Falsen E., Kersters K., Gilis M., and J. Döbereiner. 1996. Emended description of Herbaspirillum, inclusion of (Pseudomonas) rubrisubalbicans, a mild plant pathogen as Herbaspirillumrubrisubalbicans comb. nov., and classification of a group of clini- cal isolates (EF group 1) as Herbaspirillum species 3. Int. J. Syst. Bacteriol. 46: 802– 810.
Baker, A.J.M. and Walker, P.L. 1990. Ecophysiology of metal uptake by tolerant plants,
p. 155–157. In: A.J. Shaw (Ed.) Heavy metal tolerance in plants: evolutionary aspects. CRC Press Inc., Boca Raton.
Bevivino, A., Sarrocco, S., Dalmastri, C., Tabacchioni, C., Cantale, C., and Chiarini, L. 1998.
Characterization off a free-living maize-rhizosphere population of Burkholderiacepacia: effect of seed treatment on disease suppression and growth promotion of maize. FEMS Microbiology Ecology 27: 225–237.
Brewer, E.P., Saunders, J.A., Angle, J.S., Chaney, R.L., and McIntosh, M.S. 1999. Somatic
hybridization between the zinc accumulator Thlaspicaerulescens and Brassicanapus. Theo. Appl. Genet. 99: 761–771.
Burd, G.I., Dixon, D.G., and Glick, B.R. 1998. A plant growth-promoting bacterium that
decreases nickel toxicity in seedlings. Appl. Environ. Microbiol. 64: 3663–3668.
Cunningham, S.D. and Berti, W.R. 1993. Remediation of contaminated soils with green plants:
an overview. In Vitro Cell. Dev. Biol. 29: 207–212.
Cunningham, S.D. and Ow, D.W., 1996. Promises and prospects of phytoremediation. PlantPhysiol.110: 715–719. Lodewyckx et al.
Diels, L., Van Roy, S., Mergeay, M., Doyen, W., Taghavi, S., and Leysen R. 1993. Immobi-
lization of bacteria in composite membranes and development of tubular membranereactors for heavy metal recuperation. In: Effective Membrane Processes: New Perspec-tives (R. Paterson, Ed.) pp 275–293. Mechanical Engineering Publications Limited,London, UK.
Diels, L., Dong, Q., van der Lelie, D., Bayens, W., and Mergeay, M. 1995. The czc operon of
Alcaligenes eutrophus CH34: from resistance mechanism to the removal of heavy metals. J. Ind. Microbiol. 14:142–153
De Lorenzo, V., Herrero, M., Jakubzik, U., and Timmis, K.N. 1990. MiniTn5 transposon
derivatives for inserion mutagenesis, promotor probing, and chromosomal insertion of cloned DNA in Gram-negative Eubacteria. J. Bacteriol. 172: 6568–6572.
Di Fiore, S., Del Gallo, M. 1995. Endophytic bacteria: the possible role in the host plant. In:
Azospirillum VI and Related Microorganisms, Fendrik et al., Eds. Springer Verlag,Berlin, Heidelberg.
Dong, Q., Springael, D., Schoeters, J., Nuyts, G., Mergeay, M., and Diels, L. 1998. Horizontal
gene transfer of bacterial heavy metal resistance genes and its applications in activated sludge systems. Water. Sci. Tech.37: 465–468.
Figurski, D.H., Pohlman, R.F., Bechhofer, D.F., Prince, A.S., and Kelton, C.A. 1982. The
broad-host-range plasmid RK2 encodes multiple kil genes potentially lethal to Escheri- chiacoli. Proc. Natl. Acad. Sci. USA79: 1935–1939.
Hassan, M-E-T., van der Lelie, D., Springael, D., Römling, N.A., and Mergeay, M. 1999.
Identification of a gene cluster, czr, involved in cadmium and zinc resistance in Pseudomo- nasaeruginosa. Gene 238: 417–425.
Hebbar, K.P., Davey, A.G., Merrin, J., and Dart, P.J. 1992a. Rhizobacteria of maize antago-
nistic to Fusariummoniliforme, a soil borne fungal pathogen: colonization of rhizosphere and roots. Soil Biol. Biochem.24: 989–997.
Hebbar, K.P., Davey, A.G., Merrin, J., Mc Loughlin, T.J., and Dart, P.J. 1992b. Pseudomonascepacia, a potential suppressor of maize soil-borne diseases-seed inoculation and maize root colonization. Soil Biol. Biochem. 24: 999–1007.
Kloepper, J.W. and Beauchamp, C.J., 1992. A review of issues related to measuring coloni-
zation of plant roots by bacteria. Can. J. Microbiol.38: 1219–1232.
Kloepper, J.W., Schippers, B., and Bakker, P.A.H.M. 1992. Proposed elf+ination of the term
endorhizosphere. Phytopathololgy 82: 726–727.
Mehra, A. and Farago, M.E. 1994. Metal ions and plant nutrition p 32-36. In: M.E. Farago
(Ed.) Plants and Chemical Elements. Biochemistry, Uptake, Tolerance and Toxicity. VCH, Weinheim, Germany.
Mergeay, M., Nies, D., Schlegel, H.G., Gerits, J., Charles, P., and Gysegem, F.V. 1985. Alcaligenes eutrophus CH34 is a facultative chemolithotroph with plasmid-bound resis- tance to heavy metals. J. Bacteriol162: 328–334
Mergeay, M. 1997. Microbial resources for bioremediation of sites polluted by heavy metals. J.
Wild et al. (Eds.) Perspectives in Bioremediation, 65–73. Kluwer Academic Publishers.
Nies, D.H. 1992. Resistance to cadmium, cobalt, zinc and nickel in microbes. Plasmid 27: 17–
Olivares, F.L., Baldani, V.L.D., Reis, V.M., Baldani, J.I., and Döbereiner J. 1996. Occurrence
of the endophytic diazotrophs Herbaspirillum spp. in roots, stems, and leaves predomi- nantly of Gramineae. Biol. Fertil. Soils21: 197–200. Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake
Reinhold-Hurek, B. and Hurek, T. 1998. Interactions of Gramineous plants with Azoarcus spp.
and other diazotrophs: identification, localization, and perspectives to study their func-tion. Crit. Rev. Plant Sci. 17: 29-54.
Salt, D.E., Benhamou, N., Leszczyniecka, M., Raskin, I., and I. Chet. 1999. A possible role
for rhizobacteria in water treatment by plant roots. Int. J. Phytoremed. 1: 67–79.
Sambrook, J., Fritsch, E.F., and Maniatis, T. 1989. Molecular Cloning: A Laboratory Manual.
2nd ed. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
Schmidt, T. and Schlegel, H.G. 1989. nickel and cobalt resistance of various bacterial isolates
from soil and highly polluted domestic and industrial wastes. FEMS Microbiol. Ecol. 62:315-328.
Schmidt, T., Schlegel, H.G. 1994. Combined nickel-cobalt-cadmium resistance encoded by
the ncc locus of Alcaligenesxylosoxidans 31A. J Bacteriol.176: 7045–7054.
Taghavi, S., Mergeay, M., Nies, D., and van der lelie, D. 1997. Alcaligeneseutrophus as a
model system for bacterial interactions with heavy metals in the environment. Res. Microbiol. 148: 536–551.
Taghavi, S., Delanghe, H., Lodewyckx, C., Mergeay, M., and van der Lelie, D. 2001. The
nickel resistance-based minitansposons: new tool for genetic manipulation of environ- mental bacteria. Appl. Environ. Microbiol.67: 1015–1019.
van der Lelie, D., Schwuchow, T., Schwidetzky, U., Wuertz, S., Baeyens, W., Mergeay, M.,
and Nies, D. 1997. Two component regulatory system involved in transcriptional control of heavy metal homeostasis in Alcaligenes eutrophus.Mol. Microbiol. 23: 493–503.
Intravenous Immunoglobulin Therapy forStevens-Johnson SyndromeALLAN S. BRETT, MD, DYANNA PHILLIPS, and ANNETTE W. LYNN, MD, Columbia, SC ABSTRACT: Stevens-Johnson syndrome (SJS) is an acute mucocutaneous disorder that can be associated with considerable morbidity. Several previous reports, all involving either adults with acquired immunodeficiency syndrome or children , suggest that intravenous
Accreditation BRN Credits 3/10/12 Breast Cancer Risk Assessment - 7.2This activity has been planned and implemented in accordance with 3/10 - 3/11/12 National Recognition of Breast Centers of Excellence - 12.9the Essential Areas and Policies of the Accreditation Council for Continuing 3/10 - 3/11/12 Clinical Breast Examiner CertificationProgram - 7.2Medical Education (ACCME) through t
 Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake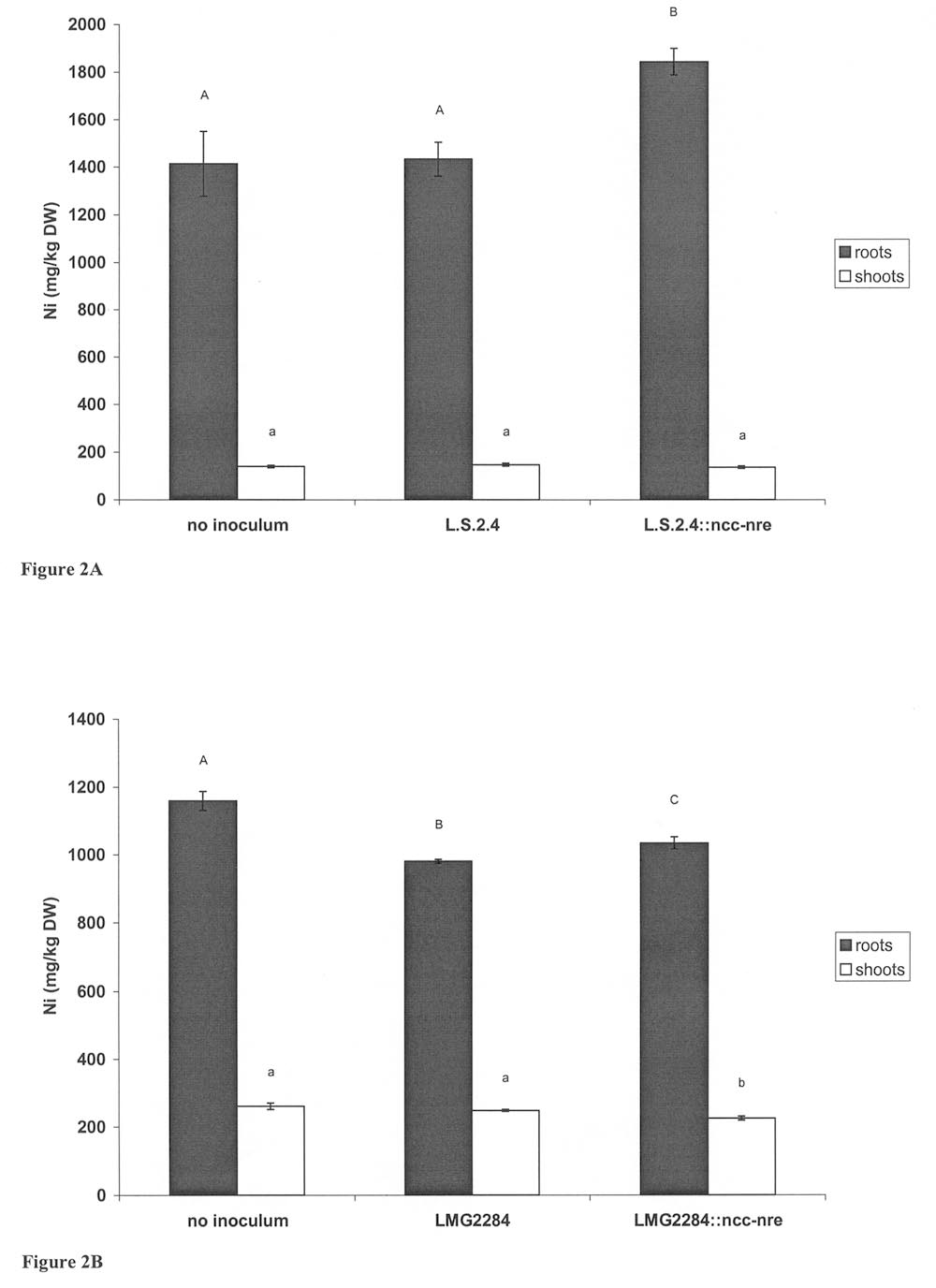 Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake
Heavy Metal-Resistant Endophytic Bacteria Effects on Heavy Metal Uptake