Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Klingberglab.se
Developmental Psychology Dopamine, Working Memory, and Training Induced Plasticity: Implications for Developmental Research Stina Söderqvist, Sissela Bergman Nutley, Myriam Peyrard-Janvid, Hans Matsson, Keith
Humphreys, Juha Kere, and Torkel Klingberg
Online First Publication, November 21, 2011. doi: 10.1037/a0026179
Söderqvist, S., Bergman Nutley, S., Peyrard-Janvid, M., Matsson, H., Humphreys, K., Kere, J.,
& Klingberg, T. (2011, November 21). Dopamine, Working Memory, and Training Induced
Plasticity: Implications for Developmental Research. Developmental Psychology. Advance
online publication. doi: 10.1037/a0026179
2011 American Psychological Association
Dopamine, Working Memory, and Training Induced Plasticity:
Stina Söderqvist, Sissela Bergman Nutley,
Karolinska Institutet and University of Helsinki
Cognitive deficits and particularly deficits in working memory (WM) capacity are common featuresin neuropsychiatric disorders. Understanding the underlying mechanisms through which WMcapacity can be improved is therefore of great importance. Several lines of research indicate thatdopamine plays an important role not only in WM function but also for improving WM capacity. Forexample, pharmacological interventions acting on the dopaminergic system, such as methylpheni-date, improve WM performance. In addition, behavioral interventions for improving WM perfor-mance in the form of intensive computerized training have recently been associated with changes indopamine receptor density. These two different means of improving WM performance—pharmacological and behavioral—are thus associated with similar biological mechanisms in thebrain involving dopaminergic systems. This article reviews some of the evidence for the role ofdopamine in WM functioning, in particular concerning the link to WM development and cognitiveplasticity. Novel data are presented showing that variation in the dopamine transporter gene (DAT1)influences improvements in WM and fluid intelligence in preschool-age children following cognitivetraining. Our results emphasize the importance of the role of dopamine in determining cognitiveplasticity. Keywords: working memory, fluid intelligence, dopamine, plasticity, cognitive training
Working memory (WM) is the ability to manipulate and keep
performance (Alloway, Gathercole, Kirkwood, & Elliott, 2009;
task relevant information in mind for a short period of time. This
Gathercole, Brown, & Pickering, 2003; Kane et al., 2004). These
is important for reasoning, which typically involves several steps
observations emphasize the importance of understanding the basis
of planning and execution. WM deficits are commonly observed in
of WM function, its development, and plasticity.
several neuropsychiatric disorders occurring during development,such as attention-deficit/hyperactivity disorder (ADHD; Castella-
Dopamine and Working Memory
nos & Tannock, 2002; Martinussen, Hayden, Hogg-Johnson, &Tannock, 2005). WM capacity is also strongly associated withgeneral intellectual ability and is a predictor for later academic
Evidence From Primates
The link between WM and dopamine has been investigated at a
cellular level by studying neurons exhibiting memory fields in theprefrontal cortex (PFC) of monkeys. These neurons are believed to
Stina Söderqvist, Sissela Bergman Nutley, and Torkel Klingberg, De-
be the cellular basis for visuospatial WM as they are specifically
partment of Neuroscience and Stockholm Brain Institute, Karolinska In-
active in response to distinct spatial locations of a stimulus and are
stitutet, Stockholm, Sweden; Myriam Peyrard-Janvid and Hans Matsson,Department of Biosciences and Nutrition at Novum, Karolinska Institutet,
also active during the delay period between stimulus presentation
Huddinge, Sweden; Keith Humphreys, Department of Medical Epidemi-
and response. Dopamine D1 receptor antagonists enhance the
ology and Biostatistics, Karolinska Institutet, Stockholm, Sweden; Juha
response of these neurons (Williams & Goldman-Rakic, 1995).
Kere, Department of Biosciences and Nutrition at Novum, Karolinska
This effect seems restricted to the D1 receptor, as no effect of a D2
Institutet, Huddinge, Sweden; Science for Life Laboratory Stockholm,
agonist was observed in the same study. Effects were also dose
Karolinska Instiutet, Stockholm, Sweden; Department of Medical Genet-
dependent and specific to the neurons displaying memory field
ics, Haartman Institute, University of Helsinki, Helsinki, Finland.
properties. D1 receptor stimulation can also lead to long-term
Torkel Klingberg and Sissela Bergman Nutley are both main inventors
improvements in WM performance, as observed in rhesus mon-
of the non-verbal reasoning training program that has been submitted in an
keys with either age-related (Castner & Goldman-Rakic, 2004) or
international patent application, financed by Pearson Assessment.
drug-induced (Castner, Williams, & Goldman-Rakic, 2000) cog-
Correspondence concerning this article should be addressed to Stina
Söderqvist, Karolinska Institutet, Department of Neuroscience, Retzius va¨g
nitive impairments. The specific mechanism by which D1 receptor
8, 171 77 Stockholm, Sweden. E-mail: stina.soderqvist@ki.se
manipulation acts to enhance visuospatial WM performance has
been suggested to occur by spatial tuning of neurons through
dopamine D1 receptor binding potential in humans 10 to 30 years
decreasing the neurons’ response to nonpreferred directions in a
of age (Jucaite, Forssberg, Karlsson, Halldin, & Farde, 2010).
spatial WM task (Vijayraghavan, Wang, Birnbaum, Williams, &
Interestingly, no age effects were observed in the posterior parietal
Arnsten, 2007). In conclusion, research on primates highlights the
cortex for the age range studied, possibly reflecting earlier devel-
importance of cortical dopamine D1 receptor activity for WM
opmental changes in this region as suggested by Goldman-Rakic
and Brown (1982). For other areas, including the frontal, anteriorcingulate, and occipital cortices, an average decrease of 26% in D1
Human Pharmacological Evidence
binding potential was observed during adolescence. This level ofdecrease is comparable with that observed throughout the entire
Evidence for dopamine’s involvement in WM functioning in
humans stems from pharmacological research, where a distinctionbetween effects of D1 and D2 receptors has also been reported. Dopamine and Development of WM Capacity
Whereas the D1/D2 receptor agonist pergolide improves perfor-mance on WM tasks, the D2 receptor agonist bromocriptine does
The effect of the gene coding for the catechol-O-
not, suggesting a predominate effect of D1 over D2 receptors for
methyltransferase (COMT) enzyme has been extensively studied
WM performance (Mu¨ller, von Cramon, & Pollmann, 1998), con-
in the context of both typical and atypical cognitive development.
sistent with the literature on primates. Further studies have dem-
The COMT enzyme is important for the degradation of cat-
onstrated pergolide’s effects to be dependent on baseline perfor-
echolamines, such as dopamine, especially within the neocortex
mance, with high-performing individuals benefiting more from the
(Matsumoto et al., 2003) and is hypothesized to be of particular
treatment than lower performers (Kimberg & D’Esposito, 2003).
importance for tasks relying on PFC functioning. The Val158Met
However, there is inconsistent evidence regarding the effect of D2
polymorphism of the COMT gene has been associated with WM
receptors and some suggestion that bromocriptine influences WM
function in both adults and children, independently and in inter-
performance in participants with low baseline WM performance
action with the DRD2 gene, coding for the dopamine D2 receptor
(Kimberg, D’Esposito, & Farah, 1997). The effect of D2 agonists
(Stelzel, Basten, Montag, Reuter, & Fiebach, 2009; Xu et al.,
on WM has also been suggested to be domain specific, improving
2007). Two studies (Barnett, Heron, Goldman, Jones, & Xu, 2009;
performance only on a spatial WM task (Luciana & Collins, 1997).
Wahlstrom et al., 2007) have observed associations between WM
Methylphenidate is a psychostimulant drug that is commonly
performance and COMT polymorphisms in children and adoles-
used to alleviate symptoms of ADHD by blocking reuptake of
cents. The effects were found to be curvilinear, with an optimal
dopamine and norepinephrine, thus increasing their availability in
level of expression being beneficial, but either too much (Val/Val
the synapse (Patrick, Caldwell, Ferris, & Breese, 1987; Solanto,
genotype) or too little (Met/Met genotype) enzymatic activity
1998). In humans, methylphenidate improves WM performance,
having negative effects on performance. Furthermore, a longitudi-
specifically in the visuospatial domain (Mehta et al., 2000). Fur-
nal study of typically developing children and adolescents dem-
thermore, the drug effect also depends on baseline WM capacity,
onstrated a developmental dependency of the COMT genotype
with greater benefits observed in participants with lower baseline
effect on WM performance (Dumontheil et al., 2011). It was
performance (Mehta, Goodyer, & Sahakian, 2004; Mehta et al.,
shown that, whereas the Val-allele tended to be associated with
superior performance on a visuospatial WM task in younger ages
In summary, human pharmacological studies provide further
(6 –10 years), the Met allele was beneficial after the age of 10. This
evidence for the importance of the dopamine neurotransmitter
is in line with the tendency for adult Met carriers to show better
system in WM function. However, the specific roles of task
WM performance than Val carriers, although evidence is currently
modality, receptor subtypes, and their interaction with subjects’
inconsistent (for a review, see Dickinson & Elvevag, 2009). These
baseline WM capacity need further investigation to provide clarity
differences in the effect of COMT observed across development
to the somewhat conflicting evidence.
are consistent with the developmental changes reported in dopa-mine concentrations discussed earlier, with changes in basal levels
Dopamine During Development
of dopamine affecting optimal levels of dopamine degradation(Wahlstrom et al., 2007).
The dopamine system is believed to undergo numerous changes
Another genetic variant that has been associated with typically
during development. In rhesus monkeys, dopamine concentration
developing WM function is a variable number tandem repeat
changes during development, with patterns of change being region
(VNTR) polymorphism located in the 3-untranslated region of the
specific (Goldman-Rakic & Brown, 1981, 1982). Lower dopamine
dopamine transporter (DAT1) gene. This polymorphism is believed
concentrations are observed in somatosensory and posterior pari-
to be involved in the expression of the gene, with higher expres-
etal cortices than in the frontal lobe. In these regions, a more rapid
sion being associated with the 10-repeat allele in vitro (Fuke et al.,
decrease in density is also observed, reaching adult levels of
2001; Mill, Asherson, Browes, D’Souza, & Craig, 2002). In chil-
concentration already at 5 months of age. In contrast, dopamine
dren ages 7 to 12 years, 9/10-repeat heterozygosity has been
levels in the prefrontal areas follow a nonlinear developmental
associated with better WM performance than 10-repeat homozy-
pattern. At birth, levels of dopamine in prefrontal areas are high
and similar to adult levels. These levels then decrease significantly
A VNTR polymorphism located in exon 3 of the dopamine
during the first 6 months of life before once again increasing to
receptor 4 gene, DRD4, has been associated with WM perfor-
reach adult levels (Goldman-Rakic & Brown, 1982). A recent
mance in young children (Froehlich et al., 2007), with carriers of
human positron emission tomography (PET) study investigated
the 7-repeat allele showing lower performance on a spatial WM
task. Furthermore the polymorphism was found to interact with the
been related to changes in activity in the caudate nucleus and
deteriorating effects of lead levels, such that children not carrying
prefrontal and parietal cortices (E. Dahlin, Neely, Larsson, Back-
the 7-repeat allele were most severely affected by lead levels and
man, & Nyberg, 2008; Olesen, Westerberg, & Klingberg, 2004).
showed worse cognitive performance. Also, activity in the PFC
For a review and discussion, see Klingberg (2010).
during performance on an N-back WM task has been associatedwith the 7-repeat allele, with young adults carrying the 7-repeat
Dopamine and Cognitive Training
allele showing a larger difference in brain activation as a result oftask difficulty (Herrmann et al., 2007). In summary, the 7-repeat is
One study has used PET to investigate the association between
associated with poorer WM performance and what can be inter-
cognitive training and dopamine D1 and D2 receptor density
preted as more ineffective brain activity. This genetic evidence
(McNab et al., 2009). Participants, who were all men in early
suggests that several genes controlling dopamine levels and sig-
adulthood, underwent a 5-week WM training scheme, training for
naling have an influential effect on WM during development.
35 min 5 days a week and were scanned using functional and
These effects on WM are likely to be complicated by Gene ϫ
structural magnetic resonance imaging and PET before and after
Gene interactions and factors affecting basal dopamine levels, such
training. The functional magnetic resonance imaging was used to
identify regions showing WM-related activity. These regionsguided PET analyses, investigating changes of dopamine D1 re-
Dopamine and Plasticity
ceptor binding potential in cortical regions and D2 receptor bind-ing potential in subcortical regions. There were no significant
Dopamine has been suggested to be important for plasticity by
associations between D1 or D2 receptor binding potential and WM
enhancing neural sprouting and synaptogenesis (Stroemer, Kent, &
performance at baseline, although a trend was observed for cortical
Hulsebosch, 1998). In stroke patients, treatment with stimulant
D1. Notably, improvements in WM capacity observed after train-
medication, which increases dopamine concentrations in the syn-
ing were significantly related to changes in D1 receptor binding
apse, enhances motor recovery resulting from physiotherapy
potential. Furthermore, fitting a quadratic model significantly in-
(Scheidtmann, Fries, Muller, & Koenig, 2001; Walker-Batson,
creased the variance explained, suggesting an inverted-U-shaped
Smith, Curtis, Unwin, & Greenlee, 1995). These effects are long
relationship, in line with previous primate and human research
lasting, with significant improvements remaining up to 12 months
findings discussed earlier. No relation between improvements in
WM capacity and D2 receptor binding potential was found. The
The effects of behavioral parenting interventions have also been
results provide further evidence for a greater importance of the D1
linked to dopaminergic function, with outcomes associated with
receptor compared with the D2 receptor not only for WM func-
variants of a VNTR in the DRD4 gene (Bakermans-Kranenburg,
tioning but also for cognitive plasticity. Note, however, the recent
Van, Pijlman, Mesman, & Juffer, 2008). This evidence is of
findings by Ba¨ckman et al. (2011), demonstrating changes in D2
particular interest as it suggests an interaction between the dopa-
receptor binding potential in the striatum after 5 weeks of updating
minergic system and environmental influences on behavioral
training. Whether the differences between these two studies result
from differences in types of training (purely updating or tasks notinvolving updating) needs further investigation. Differences might
Cognitive Training as a Model for Human
also arise from methodological differences. As McNab et al. Cognitive Plasticity
(2009) performed PET scanning during rest, changes observed inthis study reflect task-independent changes. On the other hand,
During the past decade, there has been increasing interest in
Ba¨ckman et al. (2011) used PET scanning during task performance
improving cognitive functions through targeted training. Many
and a ligand sensitive to endogenous DA release. The results might
training programs have shifted focus from explicit training, teach-
therefore also reflect changes that are task specific, such as dif-
ing strategies to improve performance, to implicit training that
ferences in behavior during scanning.
involves repetition practice, feedback, and gradually increasing the
Brehmer et al. (2009) found additional evidence for involvement
cognitive load required to solve the training tasks (Klingberg,
of the dopaminergic system in training induced plasticity from
2010). Implicit training of WM has been shown to improve per-
genetic analyses in young adults who completed a WM training
formance on nontrained WM tasks, reflecting a true increase in
program similar to the program used by McNab et al. (2009). A
WM capacity (Klingberg et al., 2005; Klingberg, Forssberg, &
sample of 29 young adults were genotyped for the DAT1 VNTR
Westerberg, 2002). Some studies report improvements in non-
and were grouped according to 10-repeat homozygosity or
trained cognitive functions associated with WM, such as attention,
9-repeat carriership (heterozygotes or homozygotes for the
reading comprehension, mathematical ability, and fluid reasoning
9-repeat allele) while controlling for the COMT Val158Met poly-
(K. Dahlin, 2011; Holmes, Gathercole, & Dunning, 2009; Jaeggi,
morphism. No significant effects of the DAT1 genotype on base-
Buschkuehl, Jonides, & Perrig, 2008; Klingberg et al., 2005;
line performance on tasks measuring WM, attention, and fluid
Klingberg et al., 2002). WM training can also have positive effects
intelligence (Gf) were observed. An initial superior (but nonsig-
on symptoms of ADHD and cognitive performance after stroke
nificant) performance on the visuospatial WM tasks used in train-
(Klingberg et al., 2005; Klingberg et al., 2002; Westerberg et al.,
ing was observed in 9/10-repeat carriers compared with 10-repeat
2007). This has obvious potential benefits for other clinical pop-
carriers. There was a trend for this difference to increase through-
ulations as well, and this area of research is currently growing
out the training period with 10-repeat carriers increasing their
rapidly. WM training is also related to changes in brain activity.
performance more steeply than noncarriers throughout the training
For example, improvements in WM observed after training have
period. No training related effects were observed for the COMT
genotype, nor were there any significant effects of either genotype
on the verbal WM tasks being trained. In summary, the effect ofthe DAT1 genotype was apparent across the training period, al-
Participants
though not seen at baseline. This pattern of results suggests that the
DNA was available from 96 children ages 4.0 to 4.5 years (56
genotype has an effect on susceptibility to training induced im-
boys and 40 girls; mean age ϭ 51.2 months, SD ϭ 3.0) who had
provements per se (E. Dahlin et al., 2008). It is important to note
completed a 5-week training scheme. Participants were recruited
that this study was underpowered for analyzing genetic effects that
with flyers distributed at preschools in the local Stockholm area
tend to be particularly small for complex traits, such as cognitive
and by advertisements in the local newspaper and on the lab
functions, and because the results were nonsignificant, a replica-
website. The children received a small reward (a toy) after com-
pletion of each set of 5 days of training. After completion of the
In the present study, we investigated the effects of polymor-
whole training period (a minimum of 20 completed training days),
phisms in five genes involved in the dopaminergic system (Table
participants received an additional monetary reward. The study
1) on the effectiveness of WM and nonverbal reasoning (NVR)
was approved by the local ethics committee, and informed consent
training in preschool children. A training program consisting of
was collected from the guardians of all participants (for further
NVR tasks was designed to assess the feasibility of improving
details, see Bergman Nutley et al., 2011).
fluid intelligence (Gf) (Bergman Nutley et al., 2011). Gf is referredto as the ability to identify patterns and relations and infer rules for
Training Program and Procedure
novel problems (Horn & Cattell, 1966). Gf is independent frompreviously learned knowledge, is highly correlated with WM ca-
Participants were pseudorandomly assigned (stratifying for sex)
pacity, and similar to WM, is a predictor of academic performance
to one of four different training programs: WM training, NVR
(Alloway & Alloway, 2010; Lynn, Meisenberg, Mikk, & Wil-
training, a CB training of WM and NVR, and a placebo training
liams, 2007). Deficits in Gf, in combination with deficits in other
designed as the CB training but with task difficulty kept constant
types of intelligence, are core symptoms of mental retardation, a
at the lowest levels throughout the training period. The WM group
particularly common mental disorder with a prevalence of around
trained with a program developed by Cogmed Systems containing
3% (Roeleveld, Zielhuis, & Gabreels, 1997). It would therefore be
seven different versions of visuospatial WM tasks. Training tasks
of great potential benefit if this function could be improved with
used for the NVR training program were based on three tests from
training similar to that used for WM. Bergman Nutley et al. (2011)
the Leiter Battery that load highly on Gf: Repeated Patterns,
recently showed that Gf could be enhanced in 4-year-old children.
Sequential Order, and Classification (Roid & Miller, 1997). Task
Each child was randomly assigned to train WM, to train NVR, or
difficulty was carefully assessed to allow for an automatic gener-
to train a combination (CB) of the two. For comparison, a placebo
ation of multiple items of varying difficulty. The WM, NVR, and
group was also included. Training of WM (in both WM and CB
CB training programs were adaptive, with level of difficulty au-
groups) resulted in significantly improved performance on WM
tomatically adjusted according to each child’s performance. Train-
transfer tasks (i.e., nontrained tasks) compared with the placebo
ing took place in the home and lasted approximately 15 min per
group. Training of NVR led to significantly improved performance
day 5 days per week until 25 sessions had been performed (Berg-
on transfer tasks of Gf, and this group also showed a trend toward
transfer between constructs with improvements on a measure of
Transfer Tests
visuospatial WM. On the basis of the evidence so far, we inves-tigated here whether these improvements were associated to poly-
Gf was measured with the Repeated Patterns, Sequential Orders,
morphisms in some genes related to the dopaminergic system.
and Classifications subtests from the Leiter Battery (Roid &
Table 1Eleven SNPs From Five Genes Involved in Dopamine-Related Pathways
a Alleles of the corresponding single nucleotide polymorphism (SNP) with the minor allele first. b Minor allele frequency (MAF) in our sample set andcorresponding allele frequency in the HapMap CEU data set in parenthesis (NA ϭ no frequency available). c Chromosomal location of the gene basedon ideogram with chromosome number and band.
d SNP position, in base pair (bp), on the respective chromosome (Genome Assembly Build 36.3).
Miller, 1997), Raven’s Coloured Progressive Matrices (Raven,
(training or no training) to assess main effect of training, and an
1998), and Block Design from the Wechsler Preschool and Pri-
interaction term of genotype and training factor to assess the effect
mary Scale of Intelligence–Third Edition (Wechsler, 2004). To
of genotype on the influence of training. Person ability was entered
assess WM capacity, we used a visuospatial grid task (Bergman
as a random effect to account for within-person correlation. The
Nutley, Söderqvist, Bryde, Humphreys, & Klingberg, 2009; West-
strongest association was found for the Gf latent variable and one
erberg, Hirvikoski, Forssberg, & Klingberg, 2004), the Odd One
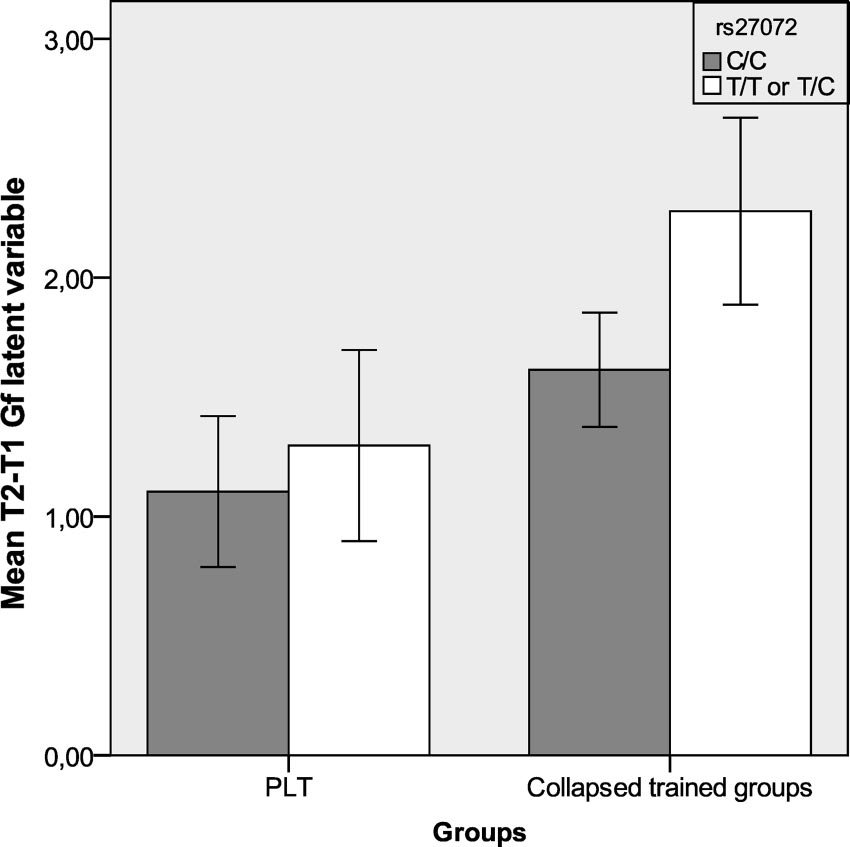
SNP (rs27072, T/C) from the DAT1 gene significantly interacting
Out task from the Automated Working Memory Assessment (Al-
with the training factor, F(4, 91) ϭ 6.971, p ϭ .01. The T-allele of
loway, 2007), and the Word Span test, a verbal WM test similar to
rs27072 seemed to be advantageous, as carriers showed a larger
the Digit Span Forward subtest from the Wechsler Intelligence
training gain than noncarriers on the Gf factor (see Figure 1). Two
Scale for Children–Third Edition (Wechsler, 1991) but with unre-
additional SNPs from the DAT1 gene also showed significant
lated nouns instead of numbers (Thorell & Wåhlstedt, 2006). For
associations. One, rs40184, was found to associate with the Gf
the measures of WM and Gf, respectively, the three measures were
latent factor, F(6, 93) ϭ 3.445, p ϭ .036, whereas another,
modeled as independent linear functions of a continuous latent
rs3863145, showed significant association with the WM latent
factor, F(6, 96) ϭ 3.545, p ϭ .032. No other polymorphismsshowed significant training interaction effects (Table 2), and no
Genes and SNPs Studied
genotype effects were observed on baseline performance. All pvalues are uncorrected for multiple comparisons of the 11 SNPs
Genetic data for the current sample were available as a subset of
tested. Thus, results should be considered preliminary and in need
a previous larger study (see Söderqvist et al., 2010). Genotypes
of replication in a larger sample set.
from single nucleotide polymorphisms (SNPs) in five genes be-longing to the dopamine pathway were available and were here
Discussion
tested for association with training performance. The genes weredopamine receptors D4 (DRD4) and D5 (DRD5), solute carrier
Previous literature suggests that dopamine might not be impor-
family 6 member 3 (SLC6A3), also known as dopamine transporter
tant only for performance on WM tasks and other cognitive tasks
(DAT1), dopamine beta-hydroxylase (DBH), and COMT. Table 1
relying on prefrontal and parietal function but also for its plasticity.
summarizes the SNPs genotyped in these genes, their chromo-
Computerized cognitive training, such as WM training (Klingberg
somal location and base pair positions according to Genome Build
et al., 2002, 2005) and reasoning training (Bergman Nutley et al.,
36.3. We selected SNP markers available from the HapMap Ge-
2011), could be useful not only for rehabilitation, but also as a
nome Browser to analyze genotype–phenotype association using
method of studying cognitive plasticity in humans, as highlighted
the genes described here. For the dopamine receptors DRD4 and
by our study. We investigated the role of genetic polymorphisms
DRD5, SNPs were selected so that they tagged the complete gene
on the change in cognitive performance resulting from training and
regions. The SNPs genotyped are in strong linkage disequilibrium
found that polymorphisms of the DAT1 gene are associated with
with the previously reported VNTR and Taq1 restriction site
training effects. As cognitive training increases in popularity, a
polymorphism for the SLC6A3/DAT1 and DBH genes, respec-tively. The VNTR markers reported in previous studies were notused for genotyping as the SNP genotyping technology availableallowed for multiplexing of a large number of SNPs in onereaction, which was the method of choice for screening a largenumber of genes. All samples had a genotyping success rate ofgreater than 95% and a genotype call rate of greater than 80%. Themethods for blood and saliva sampling, genomic DNA extraction,and SNP genotyping are described in (Söderqvist et al., 2010). Genetic Effect on Transfer
The 11 SNPs were included (separately) as fixed effects in
mixed effects models. As the dependent variable we used either thesubject loading on the latent variable for WM (Grid task, Odd OneOut task, and Word Span test) or the subject loading on the latentvariable for Gf (Leiter Battery, Raven’s Coloured ProgressiveMatrices, and Block Design tests). Because of the small number ofparticipants, all active groups (WM, NVR, and CB) were collapsedinto one training group for these analyses. The independent fixedfactors were time (Time 1, before training, or Time 2, after
Mean fluid intelligence (Gf) gains per group and genotype. Bars
training) to account for change in performance related to baseline,
show mean gains on the Gf latent variable for the collapsed trained groups and
additive effect of genotype (0, 1, or 2 copies of a specific allele of
placebo training (PLT) group and sorted by genotype on the DAT1 (rs27072)
each SNP) to assess main effect of genotype, training factor
with standard error of the mean. T1 ϭ Time 1; T2 ϭ Time 2.
ing on interactions with other genes and background factors, such
Significance Levels (p Values) From Mixed Model Analyses for
as age and baseline performance. Future studies should include
All SNPs Tested for Main Effect of Genotype on Gf and WM
larger samples to allow such interactions to be analyzed. It would
Performance as Well as Training ϫ Genotype Interactions
also be of interest to investigate the importance of domain in whichthe cognitive training is performed. As discussed earlier, there
have been suggestions that dopamine function is of particular
importance for visuospatial WM function. In the current study,
training included only visuospatial WM tasks. Thus, such a dis-
tinction is not possible. Understanding domain-specific interac-
tions with dopamine function would provide a good foundation for
better understanding how cognitive training can be individualized
to best suit people with different baseline capacities and perhaps
It has been suggested that some neurodevelopmental disorders
can be understood as disorders of learning, rather than a fixed
cognitive deficit (Karmiloff-Smith, 1998). The capacity of the
brain to adapt and learn in response to environmental influence isconsidered crucial for the development of cognitive functions.
Significant interactions indicate that training effects were associ-
Thus, a general learning impairment can lead to more specific
ated with genotype. Bold indicates p Ͻ .05 (uncorrected for multiple
disruptions later in life. The data presented here suggest that
comparisons). SNP ϭ single nucleotide polymorphism; Gf ϭ fluid intel-
dopamine is important not only for cognitive performance but
perhaps, in particular, for plasticity. The same genotype associatedwith lower plasticity in the present study is also associated with
better understanding of the mechanisms underlying its effects is of
ADHD (Gizer et al., 2009). Considering the importance of
dopamine-related genotypes in many neurodevelopmental disor-
The SNP with strongest association in our sample (rs27072) is
ders, it will be interesting to further investigate to what extent these
located in the 3Јuntranslated region of the gene and has previously
effects can be explained through an influence on learning and
been implicated in genetic studies of ADHD, with the carriers of
plasticity rather than on permanent and fixed functions.
the C/G allele exhibiting higher risk for ADHD (for a review, seeGalili-Weisstub & Segman, 2003; Gizer, Ficks, & Waldman,2009). The same allele (T) found to be advantageous in terms of
References
training gains in the present study has also been shown to have a
Alloway, T. P. (2007). Automated Working Memory Assessment. Oxford,
protective effect for ADHD (Brookes et al., 2006; Feng et al.,
2005). Furthermore, for rs27072, allelic imbalance of DAT1
Alloway, T. P., & Alloway, R. G. (2010). Investigating the predictive roles
mRNA expression has been observed, with the minor allele (T)
of working memory and IQ in academic attainment. Journal of Exper-
accounting for increased expression in both in vitro and in vivo
imental Child Psychology, 106, 20 –29. doi:10.1016/j.jecp.2009.11.003
studies (Pinsonneault et al., 2011).
Alloway, T. P., Gathercole, S. E., Kirkwood, H., & Elliott, J. (2009). The
Dopamine transporter protein removes dopamine from the syn-
cognitive and behavioral characteristics of children with low working
aptic cleft. Consequently, concentration of dopamine transporter
memory. Child Development, 80, 606 – 621. doi:10.1111/j.1467-
protein could influence the activity of both D1 and D2 receptors.
The sample size of the current study is considered small for
Ba¨ckman, L., Nyberg, L., Soveri, A., Johansson, J., Andersson, M., Dahlin,
analyses of genetic effects, and the effects observed do not remain
E., . . . Rinne, J. O. (2011, August 5). Effects of working-memorytraining on striatal dopamine release. Science, 333, 718. doi:10.1126/
significant after correcting for multiple comparisons. Thus, the
findings need further replication in larger independent samples.
Bakermans-Kranenburg, M. J., Van, I. M. H., Pijlman, F. T., Mesman, J.,
The low power is also a reason for caution when interpreting the
& Juffer, F. (2008). Experimental evidence for differential susceptibility:
lack of effects for other genotypes analyzed. It might, for example,
Dopamine D4 receptor polymorphism (DRD4 VNTR) moderates inter-
seem surprising that no effect was observed for the COMT
vention effects on toddlers’ externalizing behavior in a randomized
Val158Met polymorphism considering its extensively observed ef-
controlled trial. Developmental Psychology, 44, 293–300. doi:10.1037/
fects on WM and other cognitive functions and its importance for
degradation of dopamine in the PFC, one of the areas that show
Barnett, J. H., Heron, J., Goldman, D., Jones, P. B., & Xu, K. (2009).
training related changes in activity. It is possible that analyzing a
Effects of catechol-O-methyltransferase on normal variation in the cog-
COMT haplotype would have provided stronger effects, as has
nitive function of children. American Journal of Psychiatry, 166, 909 –
been previously suggested in studies of pain perception and atten-
Bergman Nutley, S., Söderqvist, S., Bryde, S., Humphreys, K., & Kling-
tion (Diatchenko et al., 2005) and attention alone in 2-year-old
berg, T. (2009). Measuring working memory capacity with greater
children (Voelker, Sheese, Rothbart, & Posner, 2009). Currently
precision in the lower capacity ranges. Developmental Neuropsychology,
available data do not allow for such analyses, and we suggest that
35, 81–95. doi:10.1080/87565640903325741
haplotype analysis of genetic markers in COMT can be addressed
Bergman Nutley, S., Söderqvist, S., Bryde, S., Thorell, L. B., Humphreys,
in future studies. Furthermore, effects of dopaminergic genes are
K., & Klingberg, T. (2011). Gains in fluid intelligence after training
likely to follow complicated patterns across development depend-
non-verbal reasoning in 4-year-old children: A controlled, randomized
study. Developmental Science, 14, 591– 601. doi:10.1111/j.1467-
monkeys. Neuroscience, 6, 177–187. doi:10.1016/0306-4522(81)
Brehmer, Y., Westerberg, H., Bellander, M., Furth, D., Karlsson, S., &
Goldman-Rakic, P. S., & Brown, R. M. (1982). Postnatal development of
Backman, L. (2009). Working memory plasticity modulated by dopa-
monoamine content and synthesis in the cerebral cortex of rhesus mon-
mine transporter genotype. Neuroscience Letters, 467, 117–120. doi:
keys. Developmental Brain Research, 256, 339 –349.
Herrmann, M. J., Walter, A., Schreppel, T., Ehlis, A. C., Pauli, P., Lesch,
Brookes, K., Xu, X., Chen, W., Zhou, K., Neale, B., Lowe, N., . . .
K. P., & Fallgatter, A. J. (2007). D4 receptor gene variation modulates
Johansson, L. (2006). The analysis of 51 genes in DSM–IV combined
activation of prefrontal cortex during working memory. European Jour-
type attention deficit hyperactivity disorder: Association signals in
DRD4, DAT1 and 16 other genes. Molecular Psychiatry, 11, 934 –953.
Holmes, J., Gathercole, S. E., & Dunning, D. L. (2009). Adaptive training
Castellanos, F. X., & Tannock, R. (2002). Neuroscience of attention-
leads to sustained enhancement of poor working memory in children.
deficit/hyperactivity disorder: The search for endophenotypes. NatureDevelopmental Science, 12, F9 –F15. doi:10.1111/j.1467-7687.2009
Reviews Neuroscience, 3, 617– 628. doi:10.1038/nrn896nrn896
Castner, S. A., & Goldman-Rakic, P. S. (2004). Enhancement of working
Horn, J. L., & Cattell, R. B. (1966). Refinement and test of the theory of
memory in aged monkeys by a sensitizing regimen of dopamine D1
fluid and crystallized general intelligences. Journal of Educational Psy-
receptor stimulation. Journal of Neuroscience, 24, 1446 –1450. doi:
Jaeggi, S. M., Buschkuehl, M., Jonides, J., & Perrig, W. J. (2008). Im-
Castner, S. A., Williams, G. V., & Goldman-Rakic, P. S. (2000, March 17).
proving fluid intelligence with training on working memory. Proceed-
Reversal of antipsychotic-induced working memory deficits by short-
ings of the National Academy of Sciences of the United States of
term dopamine D1 receptor stimulation. Science, 287, 2020 –2022. doi:
America, 105, 6829 – 6833. doi:10.1073/pnas.0801268105
Jucaite, A., Forssberg, H., Karlsson, P., Halldin, C., & Farde, L. (2010).
Dahlin, E., Neely, A. S., Larsson, A., Backman, L., & Nyberg, L. (2008,
Age-related reduction in dopamine D1 receptors in the human brain:
June 13). Transfer of learning after updating training mediated by the
From late childhood to adulthood, a positron emission tomography
striatum. Science, 320, 1510 –1512. doi:10.1126/science.1155466
study. Neuroscience, 167, 104 –110. doi:10.1016/j.neuroscience
Dahlin, K. (2011). Effects of working memory training on reading in
children with special needs. Reading and Writing, 24, 479 – 491. doi:
Kane, M. J., Hambrick, D. Z., Tuholski, S. W., Wilhelm, O., Payne, T. W.,
& Engle, R. W. (2004). The generality of working memory capacity: A
Diatchenko, L., Slade, G. D., Nackley, A. G., Bhalang, K., Sigurdsson, A.,
latent-variable approach to verbal and visuospatial memory span and
Belfer, I., . . . Maixner, W. (2005). Genetic basis for individual variations
reasoning. Journal of Experimental Psychology: General, 133, 189 –
in pain perception and the development of a chronic pain condition. Human Molecular Genetics, 14, 135–143. doi:10.1093/hmg/ddi013
Karmiloff-Smith, A. (1998). Development itself is the key to understanding
Dickinson, D., & Elvevag, B. (2009). Genes, cognition and brain through a COMT
developmental disorders. Trends in Cognitive Sciences, 2, 389 –398.
lens. Neuroscience, 164, 72–87. doi:10.1016/j.neuroscience.2009.05.014
Dumontheil, I., Roggeman, C., Ziermans, T., Peyrard-Janvid, M., Matsson,
Kimberg, D. Y., & D’Esposito, M. (2003). Cognitive effects of the dopa-
H., Kere, J., & Klingberg, T. (2011). Influence of the COMT genotype
mine receptor agonist pergolide. Neuropsychologia, 41, 1020 –1027.
on working memory and brain activity changes during development. Biological Psychiatry, 70, 222–229. doi:10.1016/j.biopsych
Kimberg, D. Y., D’Esposito, M., & Farah, M. J. (1997). Effects of
bromocriptine on human subjects depend on working memory capacity.
Feng, Y., Wigg, K. G., Makkar, R., Ickowicz, A., Pathare, T., Tannock,
NeuroReport, 8, 3581–3585. doi:10.1097/00001756-199711100-00032
R., . . . Barr, C. L. (2005). Sequence variation in the 3Ј-untranslated
Klingberg, T. (2010). Training and plasticity of working memory. Trends
region of the dopamine transporter gene and attention-deficit hyperac-
in Cognitive Sciences, 14, 317–324. doi:10.1016/j.tics.2010.05.002
tivity disorder (ADHD). American Journal of Medical Genetics: Neu-
Klingberg, T., Fernell, E., Olesen, P. J., Johnson, M., Gustafsson, P.,
ropsychiatric Genetics, 139B, 1– 6. doi:10.1002/ajmg.b.30190
Dahlstrom, K., . . . Westerberg, H. (2005). Computerized training of
Froehlich, T. E., Lanphear, B. P., Dietrich, K. N., Cory-Slechta, D. A.,
working memory in children with ADHD—A randomized, controlled
Wang, N., & Kahn, R. S. (2007). Interactive effects of a DRD4 poly-
trial. Journal of the American Academy of Child and Adolescent Psy-
morphism, lead, and sex on executive functions in children. Biologicalchiatry, 44, 177–186. doi:10.1097/00004583-200502000-00010
Psychiatry, 62, 243–249. doi:10.1016/j.biopsych.2006.09.039
Klingberg, T., Forssberg, H., & Westerberg, H. (2002). Training of work-
Fuke, S., Suo, S., Takahashi, N., Koike, H., Sasagawa, N., & Ishiura, S.
ing memory in children with ADHD. Journal of Clinical and Experi-
(2001). The VNTR polymorphism of the human dopamine transporter
mental Neuropsychology, 24, 781–791. doi:10.1076/jcen.24.6.781.8395
(DAT1) gene affects gene expression. Pharmacogenomics Journal, 1,
Luciana, M., & Collins, P. F. (1997). Dopaminergic modulation of working
memory for spatial but not object cues in normal humans. Journal of
Galili-Weisstub, E., & Segman, R. H. (2003). Attention deficit and hyper-
Cognitive Neuroscience, 9, 330 –347. doi:10.1162/jocn.1997.9.3.330
activity disorder: Review of genetic association studies. Israel Journal of
Lynn, R., Meisenberg, G., Mikk, J., & Williams, A. (2007). National IQs
Psychiatry and Related Sciences, 40, 57– 66.
predict differences in scholastic achievement in 67 countries. Journal of
Gathercole, S. E., Brown, L., & Pickering, S. J. (2003). Working memory
Biosocial Science, 39, 861– 874. doi:10.1017/S0021932007001964
assessments at school entry as longitudinal predictors of National Cur-
Martinussen, R., Hayden, J., Hogg-Johnson, S., & Tannock, R. (2005). A
riculum attainment levels. Educational and Child Psychology, 20, 109 –
meta-analysis of working memory impairments in children with atten-
tion-deficit/hyperactivity disorder. Journal of the American Academy of
Gizer, I. R., Ficks, C., & Waldman, I. D. (2009). Candidate gene studies of
Child and Adolescent Psychiatry, 44, 377–384. doi:10.1097/
ADHD: A meta-analytic review. Human Genetics, 126, 51–90. doi:
Matsumoto, M., Weickert, C. S., Akil, M., Lipska, B. K., Hyde, T. M.,
Goldman-Rakic, P. S., & Brown, R. M. (1981). Regional changes of
Herman, M. M., . . . Weinberger, D. R. (2003). Catechol
monoamines in cerebral cortex and subcortical structures of aging rhesus
O-methyltransferase mRNA expression in human and rat brain: Evi-
dence for a role in cortical neuronal function. Neuroscience, 116, 127–
Effects of dopamine-related gene– gene interactions on working memory
component processes. European Journal of Neuroscience, 29, 1056 –
McNab, F., Varrone, A., Farde, L., Jucaite, A., Bystritsky, P., Forssberg,
1063. doi:10.1111/j.1460-9568.2009.06647.x
H., & Klingberg, T. (2009, February 6). Changes in cortical dopamine
Stollstorff, M., Foss-Feig, J., Cook, E. H., Jr., Stein, M. A., Gaillard, W. D.,
D1 receptor binding associated with cognitive training. Science, 323,
& Vaidya, C. J. (2010). Neural response to working memory load varies
by dopamine transporter genotype in children. NeuroImage, 53, 970 –
Mehta, M. A., Goodyer, I. M., & Sahakian, B. J. (2004). Methylphenidate
977. doi:10.1016/j.neuroimage.2009.12.104
improves working memory and set-shifting in AD/HD: Relationships to
Stroemer, R. P., Kent, T. A., & Hulsebosch, C. E. (1998). Enhanced
baseline memory capacity. Journal of Child Psychology and Psychiatry
neocortical neural sprouting, synaptogenesis, and behavioral recovery
and Allied Disciplines, 45, 293–305. doi:10.1111/j.1469-7610.2004
with D-amphetamine therapy after neocortical infarction in rats. Stroke,
Mehta, M. A., Owen, A. M., Sahakian, B. J., Mavaddat, N., Pickard, J. D.,
Thorell, L. B., & Wåhlstedt, C. (2006). Executive functioning deficits in
& Robbins, T. W. (2000). Methylphenidate enhances working memory
relation to symptoms of ADHD and/or ODD in preschool children.
by modulating discrete frontal and parietal lobe regions in the human
Infant and Child Development, 15, 503–518. doi:10.1002/icd.475
brain. Journal of Neuroscience, 20, RC65.
Vijayraghavan, S., Wang, M., Birnbaum, S. G., Williams, G. V., &
Mill, J., Asherson, P., Browes, C., D’Souza, U., & Craig, I. (2002).
Arnsten, A. F. (2007). Inverted-U dopamine D1 receptor actions on
Expression of the dopamine transporter gene is regulated by the 3Ј UTR
prefrontal neurons engaged in working memory. Nature Neuroscience,
VNTR: Evidence from brain and lymphocytes using quantitative RT-
10, 376 –384. doi:10.1038/nn1846
PCR. American Journal of Medical Genetics, 114, 975–979. doi:
Voelker, P., Sheese, B. E., Rothbart, M. K., & Posner, M. I. (2009).
Variations in catechol-O-methyltransferase gene interact with parenting
Mu¨ller, U., von Cramon, D. Y., & Pollmann, S. (1998). D1- versus
to influence attention in early development. Neuroscience, 164, 121–
D2-receptor modulation of visuospatial working memory in humans.
130. doi:10.1016/j.neuroscience.2009.05.059
Journal of Neuroscience, 18, 2720 –2728.
Wahlstrom, D., White, T., Hooper, C. J., Vrshek-Schallhorn, S., Oetting,
Olesen, P. J., Westerberg, H., & Klingberg, T. (2004). Increased prefrontal
W. S., Brott, M. J., & Luciana, M. (2007). Variations in the catechol
and parietal activity after training of working memory. Nature Neuro-
O-methyltransferase polymorphism and prefrontally guided behaviors in
science, 7, 75–79. doi:10.1038/nn1165
adolescents. Biological Psychiatry, 61, 626 – 632. doi:10.1016/
Patrick, K. S., Caldwell, R. W., Ferris, R. M., & Breese, G. R. (1987).
Pharmacology of the enantiomers of threo-methylphenidate. Journal of
Walker-Batson, D., Smith, P., Curtis, S., Unwin, H., & Greenlee, R. Pharmacology and Experimental Therapeutics, 241, 152–158.
(1995). Amphetamine paired with physical therapy accelerates motor
Pinsonneault, J. K., Han, D. D., Burdick, K. E., Kataki, M., Bertolino, A.,
recovery after stroke. Further evidence. Stroke, 26, 2254 –2259. doi:
Malhotra, A. K., . . . Sadee, W. (2011). Dopamine transporter gene
variant affecting expression in human brain is associated with bipolar
Wechsler, D. (1991). Wechsler Intelligence Scale for Children–Third Edi-
disorder. Neuropsychopharmacology, 36, 1644 –1655. doi:10.1038/
tion. New York, NY: Psychological Corporation.
Wechsler, D. (2004). Wechsler Preschool and Primary Scale of
Raven, J. C. (1998). Manual for Raven’s Progressive Matrices. Oxford,
Intelligence–Third Edition (WPPSI-III). New York, NY: Psychological
United Kingdom: Oxford Psychologists Press.
Roeleveld, N., Zielhuis, G. A., & Gabreels, F. (1997). The prevalence of
Westerberg, H., Hirvikoski, T., Forssberg, H., & Klingberg, T. (2004).
mental retardation: A critical review of recent literature. Developmental
Visuo-spatial working memory span: A sensitive measure of cognitive
Medicine and Child Neurology, 39, 125–132. doi:10.1111/j.1469-
deficits in children with ADHD. Child Neuropsychology, 10, 155–161.
Roid, G. H., & Miller, L. J. (1997). Leiter International Performance
Westerberg, H., Jacobaeus, H., Hirvikoski, T., Clevberger, P., Ostensson,
Scale–Revised: Examiner’s manual. Wood Dale, IL: Stoelting.
M. L., Bartfai, A., & Klingberg, T. (2007). Computerized working
Scheidtmann, K., Fries, W., Muller, F., & Koenig, E. (2001). Effect of
memory training after stroke—A pilot study. Brain Injury, 21, 21–29.
levodopa in combination with physiotherapy on functional motor recov-
ery after stroke: A prospective, randomised, double-blind study. Lancet,
Williams, G. V., & Goldman-Rakic, P. S. (1995, August 17). Modulation
358, 787–790. doi:10.1016/S0140-6736(01)05966-9
of memory fields by dopamine D1 receptors in prefrontal cortex. Nature,
Söderqvist, S., McNab, F., Peyrard-Janvid, M., Matsson, H., Humphreys,
376, 572–575. doi:10.1038/376572a0
K., Kere, J., & Klingberg, T. (2010). The SNAP25 gene is linked to
Xu, H., Kellendonk, C. B., Simpson, E. H., Keilp, J. G., Bruder, G. E.,
working memory capacity and maturation of the posterior cingulate
Polan, H. J., . . . Gilliam, T. C. (2007). DRD2 C957T polymorphism
cortex during childhood. Biological Psychiatry, 68, 1120 –1125. doi:
interacts with the COMT Val158Met polymorphism in human working
memory ability. Schizophrenia Research, 90, 104 –107. doi:10.1016/
Solanto, M. V. (1998). Neuropsychopharmacological mechanisms of stim-
ulant drug action in attention-deficit hyperactivity disorder: A reviewand integration. Behavioural Brain Research, 94, 127–152. doi:10.1016/
Stelzel, C., Basten, U., Montag, C., Reuter, M., & Fiebach, C. J. (2009).
Sicherheitsdatenblatt gemäß 91/155/EWG 1 Bezeichnung des Stoffes/der Zubereitung und des Unternehmens · Angaben zum Produkt · Handelsname: RUCOZID Holzwurmbekämpfung · Artikelnummer: OP251662, 88251662 · Verwendung des Stoffes / der Zubereitung Holzschutzmittel/Anwendung gemäss technischem Merkblatt · Hersteller/Lieferant: Rupf & Co AGFarben- u.
UM ESTUDO SUPLEMENTAR A VELHA LEI NO NOVO TESTAMENTO OWEN D. OLBRICHT “Agora, com efeito, obteve Jesus ministério tan- digo: até que o céu e a terra passem, nem um i ou to mais excelente, quanto é ele também Mediador um til jamais passará da Lei, até que tudo se de superior aliança instituída com base em supe- riores promessas. Porque, se aquela primeira Essa passage
 Miller, 1997), Raven’s Coloured Progressive Matrices (Raven,
(training or no training) to assess main effect of training, and an
1998), and Block Design from the Wechsler Preschool and Pri-
interaction term of genotype and training factor to assess the effect
mary Scale of Intelligence–Third Edition (Wechsler, 2004). To
of genotype on the influence of training. Person ability was entered
assess WM capacity, we used a visuospatial grid task (Bergman
as a random effect to account for within-person correlation. The
Nutley, Söderqvist, Bryde, Humphreys, & Klingberg, 2009; West-
strongest association was found for the Gf latent variable and one
erberg, Hirvikoski, Forssberg, & Klingberg, 2004), the Odd One
SNP (rs27072, T/C) from the DAT1 gene significantly interacting
Out task from the Automated Working Memory Assessment (Al-
with the training factor, F(4, 91) ϭ 6.971, p ϭ .01. The T-allele of
loway, 2007), and the Word Span test, a verbal WM test similar to
rs27072 seemed to be advantageous, as carriers showed a larger
the Digit Span Forward subtest from the Wechsler Intelligence
training gain than noncarriers on the Gf factor (see Figure 1). Two
Scale for Children–Third Edition (Wechsler, 1991) but with unre-
additional SNPs from the DAT1 gene also showed significant
lated nouns instead of numbers (Thorell & Wåhlstedt, 2006). For
associations. One, rs40184, was found to associate with the Gf
the measures of WM and Gf, respectively, the three measures were
latent factor, F(6, 93) ϭ 3.445, p ϭ .036, whereas another,
modeled as independent linear functions of a continuous latent
rs3863145, showed significant association with the WM latent
factor, F(6, 96) ϭ 3.545, p ϭ .032. No other polymorphismsshowed significant training interaction effects (Table 2), and no
Genes and SNPs Studied
Miller, 1997), Raven’s Coloured Progressive Matrices (Raven,
(training or no training) to assess main effect of training, and an
1998), and Block Design from the Wechsler Preschool and Pri-
interaction term of genotype and training factor to assess the effect
mary Scale of Intelligence–Third Edition (Wechsler, 2004). To
of genotype on the influence of training. Person ability was entered
assess WM capacity, we used a visuospatial grid task (Bergman
as a random effect to account for within-person correlation. The
Nutley, Söderqvist, Bryde, Humphreys, & Klingberg, 2009; West-
strongest association was found for the Gf latent variable and one
erberg, Hirvikoski, Forssberg, & Klingberg, 2004), the Odd One
SNP (rs27072, T/C) from the DAT1 gene significantly interacting
Out task from the Automated Working Memory Assessment (Al-
with the training factor, F(4, 91) ϭ 6.971, p ϭ .01. The T-allele of
loway, 2007), and the Word Span test, a verbal WM test similar to
rs27072 seemed to be advantageous, as carriers showed a larger
the Digit Span Forward subtest from the Wechsler Intelligence
training gain than noncarriers on the Gf factor (see Figure 1). Two
Scale for Children–Third Edition (Wechsler, 1991) but with unre-
additional SNPs from the DAT1 gene also showed significant
lated nouns instead of numbers (Thorell & Wåhlstedt, 2006). For
associations. One, rs40184, was found to associate with the Gf
the measures of WM and Gf, respectively, the three measures were
latent factor, F(6, 93) ϭ 3.445, p ϭ .036, whereas another,
modeled as independent linear functions of a continuous latent
rs3863145, showed significant association with the WM latent
factor, F(6, 96) ϭ 3.545, p ϭ .032. No other polymorphismsshowed significant training interaction effects (Table 2), and no
Genes and SNPs Studied