Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Pii: s0021-9924(01)00060-0
Kenneth R. Pugha,b,*, W. Einar Mencla,b, Annette R. Jennera,b,
Leonard Katzb,c, Stephen J. Frostb,c, Jun Ren Leea,b,
Sally E. Shaywitza, Bennett A. Shaywitza,d
aDepartment of Pediatrics, Yale University School of Medicine, PO Box 3333, New Haven,
bHaskins Laboratories, New Haven, CT 06511, USA
cDepartment of Psychology, University of Connecticut, Storrs, CT, USA
dDepartment of Neurology, Yale University School of Medicine, New Haven CT, USA
Received 4 May 2001; received in revised form 25 June 2001; accepted 25 June 2001
Evidence from neuroimaging studies, including our own, suggest that skilled word
identification in reading is related to the functional integrity of two consolidated lefthemisphere (LH) posterior systems: a dorsal (temporo-parietal) circuit and a ventral(occipito-temporal) circuit. This posterior system appears to be functionally disrupted indevelopmental dyslexia. Relative to nonimpaired readers, reading-disabled individualsdemonstrate heightened reliance on both inferior frontal and right hemisphere posteriorregions, presumably in compensation for the LH posterior difficulties. We propose aneurobiological account suggesting that for normally developing readers, the dorsal circuitpredominates at first, and in conjunction with premotor systems, is associated with analyticprocessing necessary for learning to integrate orthographic with phonological and lexical –semantic features of printed words. The ventral circuit constitutes a fast, late-developing,word form system, which underlies fluency in word recognition. Learning outcomes: As a result of this activity, (1) the participant will learn about amodel of lexical processing involving specific cortical regions. (2) The participant willlearn about evidence which supports the theory that two dorsal LH systems may bedisrupted in developmental dyslexia. (3) The participant will learn that individuals with
* Corresponding author. Haskins Laboratories, 270 Crown Street, New Haven, CT 06511, USA.
Tel.: +1-203-865-6163x224; fax: +1-203-764-9353.
E-mail address: pugh@tom.haskins.yale.edu (K.R. Pugh).
0021-9924/01/$ – see front matter D 2001 Elsevier Science Inc. All rights reserved. PII: S 0 0 2 1 - 9 9 2 4 ( 0 1 ) 0 0 0 6 0 - 0
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
reading impairment may rely on other regions of the brain to compensate for the disruptionof posterior function. D 2001 Elsevier Science Inc. All rights reserved.
Keywords: Dyslexia; Neuroimaging; Reading; Lexical processing; Phonology
Theories of reading disability have been proposed based on putative
deficiencies of several kinds of processing: the visual system (Stein & Talcott,1999; Stein, 1993), the language system (Shankweiler, Liberman, Mark,Fowler, & Fischer, 1979), and the fine-grained temporal processing of auditorystimuli (Stein & Walsh, 1997; Tallal & Stark, 1982). However, there is now agood consensus among reading researchers that whatever the contributions ofother systems and processes may be, the core difficulty in reading disabilitymanifests itself as a deficiency within the language system and, in particular, adeficiency at the level of phonological analysis. To learn to read, a child mustfirst develop an appreciation of the segmental nature of speech and come torealize that spoken words are composed of the smallest of these segments —the phoneme. This appreciation of the segmental nature of speech is termedphonemic awareness. Subsequently, the beginning reader must also understandthat written words, too, possess an internal phonological structure that is thesame as the spoken word. It is phonemic awareness and the understanding thatthe constituents of a printed word bear a relationship to phonemes that allowsthe reader to connect printed words to the corresponding words in his/herspeech lexicon.
As many studies have shown, phonemic awareness is largely missing in
reading-disabled (RD) children and adults (Brady & Shankweiler, 1991; Bruck,1992; Fletcher et al., 1994; Rieben & Perfetti, 1991; Shankweiler et al., 1995;Shaywitz et al., 1999; Stanovich & Siegel, 1994). As to why RD readers shouldhave exceptional difficulty developing phonemic awareness, there is support forthe notion that the difficulty resides in the phonological component of the largerspecialization for spoken language (Liberman, 1998; Liberman, Shankweiler, &Liberman, 1989). If that component is imperfect in an individual, the individ-ual’s perception of phonemes will be less than ideally distinctive. Therefore, itwill be harder to bring their distinctiveness to conscious awareness, as must bedone when learning the ‘‘sounds that letters make,’’ i.e., in memorizing thecorrespondences between letters and letter clusters, on one hand, and thephonemes and syllables that they represent, on the other. As noted above,there is now overwhelming evidence that phonological awareness is character-istically deficient (or lacking) in RD readers who, as a consequence, havedifficulty mapping the alphabetic characters of print onto the spoken word. Forexample, measures of phonemic awareness predict later reading achievement
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
(Bradley & Bryant, 1983; Stanovich, Cunningham, & Cramer, 1984); deficits inphonemic awareness consistently separate RD and nondisabled children(Fletcher et al., 1994; Stanovich & Siegel, 1994); phonological processingdeficits persist into adulthood (Bruck, 1992; Felton, Naylor, & Wood, 1990;Shaywitz et al., 1999) and instruction in phonemic awareness promotes theacquisition of reading skills (Ball & Blachman, 1991; Bradley & Bryant, 1983;Foorman, Francis, Fletcher, Schatschneider, & Mehta, 1998; Torgesen, Morgan,& Davis, 1992; Wise & Olson, 1995). Given this background, our ownfunctional neuroimaging research program and studies selected for discussionin this review involve a comparison of RD and nonimpaired (NI) readinggroups on word and pseudoword reading tasks that stress phonologicalprocessing. For a discussion of functional neuroimaging studies that haveexamined sensory-level processing deficits in developmental dyslexia (e.g.,Demb, Boynton, & Heeger, 1998; Eden et al., 1996), the reader is referred toEden and Zeffiro (1998). For reviews of research examining anatomical/structural differences between RD and NI groups, the reader is referred toFilipek (1995) and Galaburda (1992).
2. Neurobiological studies: the posterior and anterior reading circuits
There is substantial converging evidence that identification of printed words
implicates a posterior cortical reading system with both ventral and dorsalcomponents. The ventral circuit includes lateral extrastriate areas and a leftinferior occipito-temporal area where functional imaging studies show robustactivation in word-reading tasks, and where electrophysiological studies revealthe earliest source of temporal dissociation (approximately 150 – 180 ms) insignal between printed words and pseudowords, on one hand, and nonlinguisticvisual materials, on the other (Fiez & Petersen, 1998; Frackowiak, Friston, Frith,Dolan, & Mazziotta, 1997; Henderson, 1986; see also Nobre, Allison, &McCarthy, 1994; Puce, Allison, Asgari, Gore, & McCarthy, 1996; Salmelin,Service, Kiesila, Uutela, & Salonen, 1996; Tarkiainen, Helenius, Hansen,Cornelisssen, & Salmelin, 1999). Evidence from a number of functional imagingstudies finds this circuit to be disrupted in RD individuals (Brunswick, McCrory,Price, Frith, & Frith, 1999; Helenius, Tarkiainen, Cornelissen, Hansen, &Salmelin, 1999; Helenius, Uutela, & Hari, 1999; Pugh et al., 2000; Rumsey etal., 1997; Salmelin et al., 1996; Shaywitz et al., 1998, submitted).
A more dorsal reading-related system contains the angular gyrus and supra-
marginal gyrus in the inferior parietal lobule, and the posterior aspect of thesuperior temporal gyrus (Wernicke’s Area). The temporo-parietal circuit has longbeen implicated in reading and writing disorders. Indeed, a large literature onacquired dyslexia indicates that lesions centered about the angular gyrus arestrongly predictive of this malady (Damasio & Damasio, 1983; Dejerine, 1891;Friedman, Ween, & Albert, 1993; Henderson, 1986). On several accounts, this
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
region is considered relevant in mapping the visual percepts of print onto thephonological structures of language (Benson, 1994; Black & Behrmann, 1994;Geschwind, 1965). Converging findings from functional imaging studies alsoimplicate the temporo-parietal circuit in RD, indicating abnormal activationduring reading-related tasks when decoding and when other linguistic analysesare taxed (Flowers, Wood, & Naylor, 1991; Gross-Glenn et al., 1991; Horwitz,Rumsey, & Donohue, 1998; Pugh et al., 2000; Rumsey et al., 1992, 1997;Salmelin et al., 1996; Shaywitz et al., 1998, submitted; Simos et al., 2000).
Anterior sites centered in and around Broca’s Area in the inferior frontal gyrus
(IFG) appears to be associated with fine-grained, speech – gestural (articulatory),phonological recoding. This circuit also functions in silent reading and naming(reviewed in Fiez & Petersen, 1998; Frackowiak et al., 1997; Pugh et al., 1996,1997). Functional imaging studies implicate this inferior frontal region in RD(Brunswick et al., 1999; Paulesu et al., 1996; Rumsey et al., 1997; Shaywitz etal., 1998, submitted). Several findings suggest that the anterior system appears tobe more heavily used by RD than normal readers, perhaps in compensation fortheir failure to develop the posterior reading system adequately (Pugh et al.,2000; Shaywitz et al., 1998).
3. Functional roles for posterior and anterior circuits in NI
Neuroimaging studies reveal several important differences between the
temporo-parietal (dorsal) and occipito-temporal (ventral) left hemisphere (LH)posterior regions, which allow speculation on their distinctive roles in skilledword and pseudoword reading. From these findings, we suggest that the temporo-parietal circuit is associated with slow decoding, i.e., a rule-based analysis of theprinted word that makes heavy use of attentional resources. This system is criticalfor extracting and learning the relationships between orthography and itsphonological forms, connecting these to morphological and lexical – semanticinformation. With repeated experience of a particular word, all these elementsbecome bound into highly integrated representations. Thus, basic decoding andanalysis skills rely on an intact temporo-parietal organization.
The ventral circuit lies at the point of contact between the ventral visual stream
and the middle to inferior temporal lobe. Our cross-sectional study of childrenindicates that the ventral word form area plays an increasingly important role inword identification for young NI readers as they become skilled (Shaywitz et al.,submitted). In contrast to the dorsal circuit, the ventral occipito-temporal arearesponds rapidly in word identification and may, we conjecture, respondautomatically, without heavy dependence on attentional resources. We suggestthat the information coded by the ventral circuit is linguistically structuredorthographic form. Note, however, that although the ventral system appears toprocess information that is orthographic (i.e., in the visual modality), we proposethat this information is structured perceptually in linguistic terms, i.e., in
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
orthographic units that correspond to the phonological and morphological unitsof the speech form of the word. Such sensitivity to phonological and morpho-logical structure could be the legacy of the slower dorsal circuit decoding process,the system that initially learned how to recognize a particular printed word. Thus,we propose that the earlier developing dorsal circuit guides and shapes thedevelopment of the ventral circuit. The development of the ventral word formcircuit may depend on the integrity of the word-analytic temporo-parietal (dorsal)circuit that precedes it.
What is the evidence that supports this distinction between the dorsal and
ventral circuits? In skilled readers, the dorsal circuit responds with greateractivation to pseudowords and low-frequency words (words that must bedecoded effortfully) than to familiar high-frequency words (see Frackowiak etal., 1997 for discussion). The ventral system shows the opposite response profilewith higher activation to familiar words than to pseudowords (Tagamets,Novick, Chalmers, & Friedman, 2000; see Frackowiak et al., 1997 for a reviewof evidence from positron emission tomography (PET) studies regarding thesesystems). The increased response to unfamiliar stimuli by the dorsal systemsuggests that it is engaging in decoding, an effortful phonological analysis. Bycontrast, the ventral circuit appears to support a type of processing that dependsstrongly on familiarity (i.e., memory): activation is higher for well-learnedwords. Further evidence that suggests this distinction shows that simple wordidentification tasks make maximum demands on ventral sites with limiteddemands on dorsal sites (Brunswick et al., 1999), while tasks that involvephonological or semantic analysis show heightened dorsal responses (Pugh et al.,1996, 2000; Rumsey et al., 1997; Shaywitz et al., 1998). Additionally, Price,Moore, & Frackowiak (1996) demonstrated that as stimulus presentation ratesincreased from 20 through 60 words/min, ventral areas showed increasedactivation while dorsal areas showed decreased activation, again an oppositeprofile. It is plausible that analysis and computation for a given stimulus areprecluded at very fast input rates, but basic visual pattern identification processesare intact. This is consistent with the notion that the dorsal circuit maps letters tophonology via effortful decoding (a computational or algorithmic process) whilethe ventral circuit is associated with rapid stimulus identification in which astimulus that is perceived as a structured pattern is matched to a similar patternin memory. Finally, the ventral circuit responds more rapidly. As noted above,Salmelin and her colleagues (Salmelin et al., 1996; Tarkiainen et al., 1999),using MEG, demonstrated that evoked responses to words and pseudowordsdiverge from nonlinguistic visual stimuli early (between 150 and 180 ms) at theoccipito-temporal area; temporo-parietal responses arise later in time (approx-imately 250 ms). Further, this difference occurred in skilled readers but not inRD readers.
With respect to the anterior circuit including the LH IFG, studies indicate that
it is active in silent reading (Fiez & Petersen, 1998; Price et al., 1996; Pugh et al.,1996, 1997), and is more strongly engaged by low-frequency words and
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
pseudowords than by high-frequency words; in this regard, it mirrors the dorsalcircuit (Fiez & Petersen, 1998). A number of findings suggest that the anteriorsystem has a role in the speech – gestural (articulatory) recoding of print. Forexample, we observed significantly higher levels of activation in IFG duringword- and pseudoword-naming than during silent reading of the same stimuli —another indication of a role for this anterior circuit in gestural phonologicalrecoding (Pugh et al., in preparation). Further, the circuit shows a high degree ofsensitivity to the spelling – sound regularity (or perhaps consistency) of words. Pugh et al. (1997) found that interhemispheric variation in IFG activity predictedindividuals’ sensitivities to regularity effects in silent reading tasks. Low-frequency, irregularly spelled words (e.g., PINT) elicited higher activation thanregular words (e.g., MINT) at this site (Herbster, Mintun, Nebes, & Becker,1997; see also Fiez & Petersen, 1998), a finding that converges with those ofPugh et al. (1997) in suggesting a relation between the IFG circuit and regularity/consistency effects. Behavioral studies have shown that, on average, regularity/consistency effects are stronger in overt speech production tasks (outputphonology) than in silent reading tasks such as lexical decision, suggesting animportant contribution of gestural phonological recoding to this effect (Hino &Lupker, 2000). Indeed, several studies have shown significant effects in delayednaming tasks, suggesting that at least part of the effect is related to articulatoryrecoding in overt production tasks (Inhoff, Briihl, & Schwartz, 1996; Ziegler,Montant, & Jacobs, 1997).
4. Altered functions for posterior and anterior circuits in RD
There are clear functional differences between NI and RD readers with regard
to the dorsal, ventral, and anterior sites we have been discussing. In RD readers,a number of functional imaging studies have observed LH posterior dysfunction,at both dorsal and ventral sites during phonological processing tasks (Brunswicket al., 1999; Helenius, Tarkiainen, et al., 1999; Helenius, Uutela, et al., 1999;Pugh et al., 2000; Rumsey et al., 1992, 1997; Salmelin et al., 1996; Shaywitz etal., 1998, submitted; Simos et al., 2000). This disruption is reflected by a relativeunderengagement of these circuits specifically in processing words and pseudo-words where decoding is required, suggesting a disruption of this region in RDreaders. For instance, in our study of adults (Shaywitz et al., 1998), we observeddifferences between RD and NI readers in the patterns of activation in severalcritical components of the LH posterior reading system: posterior STG (Wer-nicke’s Area), angular gyrus, occipito-temporal areas, and striate cortex. Thepattern of group differences was similar at each of these sites: NI readers showeda systematic increase in activation as orthographic-to-phonologic processingdemands increased, while RD readers failed to show such systematic modulationin their activation patterns in response to the same task demands. As notedabove, in the Salmelin studies using MEG, skilled readers show a response to
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
printed tokens between 150 and 180 ms poststimulus onset at the occipito-temporal area, but this early ventral response is not seen in adult developmentaldyslexics, suggesting disruption here for them. Additionally, in our recentexamination of children (Shaywitz et al., submitted), we observed anomalousactivation in RD readers at both dorsal and ventral LH sites during word- andpseudoword-reading tasks, confirming our findings from an earlier adult sample(Shaywitz et al., 1998). Importantly, this posterior anomaly was confined to tasksthat tapped word- and pseudoword-reading only, for both adults and children. Incontrast, on tasks that did not require phonological decoding, no group differ-ences were seen.
While most neuroimaging studies have sought to identify those brain regions
within which activation patterns discriminate RD from NI readers, implicatingthe dorsal and ventral aspects of LH posterior cortex, a more completeunderstanding of the neurobiology of developmental dyslexia requires that wealso consider relations between the distinct brain regions, which functioncooperatively to process information during reading. This relational processingissue has been referred to as one of functional connectivity between corticalareas. Evidence consistent with the notion of a breakdown in functionalconnectivity within the posterior reading system in RD readers has been recentlyreported by Horwitz et al. (1998) using activation data from the Rumsey et al. (1997) PET study. These authors examined correlations between activationlevels in the LH angular gyrus and other brain sites during two reading-aloudtasks. Correlations between the LH angular gyrus and occipital and temporallobe sites were strong and significant in NI readers and weak in RD readers. Such a result suggests a breakdown in functional connectivity across the majorcomponents of the posterior reading system. We recently extended the analysisof our initial sample of adults to examine functional connectivity between LHposterior regions in these two groups (Pugh et al., 2000). As in the Horwitz et al. study, we looked at functional connectivity between the angular gyrus andoccipital and temporal lobe sites, but using a hierarchically organized set oftasks — tasks that systematically varied demands made on phonologicalassembly. While for RD readers LH functional connectivity was indeed weakon word- and nonword-reading tasks as suggested by Horwitz et al., thereappeared to be no dysfunction in the tasks which tap metaphonological judg-ments only, or complex visual – orthographic coding only. The results are mostconsistent with a specific phonological deficit hypothesis: A breakdown in LHposterior systems manifests only when orthographic to phonological assembly isrequired. Moreover, we found that on word- and nonword-reading tasks, righthemisphere (RH) homologues appear to function in a compensatory manner forRD readers; correlations were strong and stable in this hemisphere for bothreading groups.
In our studies (Pugh et al., 2000; Shaywitz et al., 1998, submitted), we also
observed reading disability processing that appears to be compensatory. Wefound that on those tasks that made explicit demands on phonological
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
processing (pseudoword and word tasks), RD readers showed a disproportion-ately greater engagement of IFG and prefrontal dorsolateral sites than did NIreaders. Rumsey et al. (1997) found the same pattern of posterior disruptionand inferior frontal reliance with PET. As noted, a MEG study by Salmelin et al. (1996) found evidence of a relative early frontal response in RD readerscoupled with the occipito-temporal anomaly discussed previously and, morerecently, Brunswick et al. (1999) and Richards et al. (1999) have showndisproportionately elevated frontal activation in RD readers across severaldifferent tasks. Thus fMRI, PET, and MEG studies coverage to suggest botha LH posterior anomaly and an apparent compensatory shift to frontal sites inreading disability.
Evidence of a second apparent compensatory shift (in this case, to posterior
RH), comes from several findings. In Shaywitz et al. (1998), we observed asignificant interaction of hemisphere by reading group in angular gyrus andmiddle temporal gyrus. The effect showed greater RH, than LH, activation in RDreaders, but greater LH, than RH, activation in NI readers (see also Barnea,Lamm, Epstein, & Pratt, 1994). As noted, in the correlational analysis with thesesame reading tasks, RD readers failed to demonstrate any evidence of functionalconnectivity between major posterior circuits in the LH. In contrast, theydisplayed strong correlations at reading disability homologues of these sites thatwere numerically higher than the correlations of NI readers (Pugh et al., 2000). Rumsey et al. (1999) examined the relationship between RH activation andreading performance in their RD and NI subjects and found that RH temporo-parietal activation was correlated with standard measures of reading performanceonly for RD readers, suggesting a compensatory function for the shift to RH. Weobserved a similar predictive relation between RH sites and reading skill in oursamples of children (Shaywitz et al., submitted). In summary, NI readers show astrong functioning LH posterior circuit in word- and pseudoword-reading, butRD readers do not. Instead, they show evidence of two, apparently compensatory,responses to their LH posterior dysfunction: increased bi-hemispheric IFGactivation and an increased functional role for RH posterior sites (Pugh et al.,2000; Shaywitz et al., 1998).
Posterior reading circuits including both dorsal (temporo-parietal) and ventral
(occipito-temporal) components are disrupted in people who are RD, as indicatedby reduced activation as well as by disrupted functional connectivity betweenthese areas. Additionally, there appear to be two characteristic compensatorypatterns in response to this LH posterior anomaly: (1) increased reliance on IFGduring reading, and (2) an increased tendency to rely on the RH homologues ofthe dysfunctional LH posterior circuits. Our cross-sectional imaging studies ofchildren suggest the following model.
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
In the normally developing NI reader, the development of the LH posterior
reading circuits, particularly the ventral occipito-temporal area, is dependent onan organized integration of phonological, morphological, and lexical – semanticprocessing of words within highly overlapping neural circuits. We assume thatthis integration relies initially on the intactness of processing in the temporo-parietal circuit (presumably via feedforward and feedback resonance witharticulatory recoding sites in the frontal lobe). Deficient dorsal function willfail to support appropriate ventral development. Thus, in the RD reader,temporo-parietal difficulties disrupt this developmental trajectory. The shift toinferior frontal sites in the RD child reflects a compensatory reliance on thesecircuits to support articulatory recoding (covert pronunciation) in an attempt tocope with a problematic phonological analysis of printed words. A secondcompensatory shift, from posterior LH to posterior RH, likely reflects thedevelopment of an additional word recognition process that is essentiallyvisual – perceptual; graphemic patterns in the printed word are associateddirectly with entries in the RD reader’s mental lexicon. Thus, these visualpatterns do not code the phonological or morphological information that the NIreader perceives within the printed word, but instead represent the printed wordas a nonlinguistic visuo-semantic icon. This developmental account of theneural circuitry of reading (Fig. 1) provides a first approximation for theneurobiological substrate of reading and RD. As the model develops, we canbegin to provide a means for the evaluation and monitoring of interventions andreading remediation programs. For example, investigators might choose to focus
Fig. 1. A tentative model of the neural circuitry for reading.
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
on training-related changes in the activation of the LH ventral circuit as a targetfor interventions that could be expected in turn to lead to improved word-reading skill in dyslexic children.
The writing of this manuscript was supported by grants from the National
Institute of Child Health and Human Development (PO1 HD-21888 and P50 HD-2580) to Yale School of Medicine and by National Institute of Child Health andHuman Development grant HD-01994 to Haskins Laboratories. This paper isdedicated to the memory of our dear friend and colleague, Professor AlvinLiberman, whose intellectual guidance and boundless love of truth was, and willcontinue to be, a driving force behind our program of research.
1. Reading impairment in the majority of individuals with dyslexia appears
a. visual confusionsb. letter confusionsc. phonological decoding problemsd. vocabulary deficitse. syntactic deficits
2. The posterior ventral reading circuit appears to support
a. identification of printed wordsb. identification of sentence contextc. identification of sword meaningsd. phonological decodinge. syntactic decoding
3. Reading-disabled subjects may attempt to compensate for poor
a. increased reliance of the ventral posterior regions of the brainb. increased reliance on the dorsal posterior regions of the brainc. increased reliance on the superior frontal regions of the braind. increased reliance on the inferior frontal regions of the braine. increased reliance on the RH
4. The dorsal reading circuit appears to support
a. phonological analysisb. syntactic analysis
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
c. morphological analysisd. high-frequency wordse. all of the above
5. The inferior frontal region of the brain tends to be active in tasks requiring
a. decoding of pseudowordsb. decoding of low-frequency wordsc. articulatory recoding of printd. silent readinge. nonreading control tasks
Ball, E. W., & Blachman, B. A. (1991). Does phoneme awareness training in kindergarten make a
difference in early word recognition and developmental spelling? Reading Research Quarterly, 26,49 – 66.
Barnea, A., Lamm, O., Epstein, R., & Pratt, H. (1994). Brain potentials from dyslexic children
recorded during short-term memory tasks. International Journal of Neuroscience, 74, 227 – 237.
Benson, D. F. (1994). The neurology of thinking. New York: Oxford University Press. Black, S. E., & Behrmann, M. (1994). Localization in alexia. In: A. Kertesz (Ed.), Localization and
neuroimaging in neuropsychology ( pp. 331 – 376). New York: Academic Press.
Bradley, L., & Bryant, P. E. (1983). Categorizing sounds and learning to read — a causal connection.
Brady, S. A., & Shankweiler, D. P. (Eds.) (1991). Phonological processes in literacy: a tribute to
Isabelle Y. Liberman. Hillsdale, NJ, USA: Lawrence Erlbaum Associates.
Bruck, M. (1992). Persistence of dyslexics’ phonological deficits. Developmental Psychology, 28,
Brunswick, N., McCrory, E., Price, C., Frith, C. D., & Frith, U. (1999). Explicit and implicit process-
ing of words and pseudowords by adult developmental dyslexics: a search for Wernicke’s Wort-schatz? Brain, 122, 1901 – 1917.
Damasio, A. R., & Damasio, H. (1983). The anatomic basis of pure alexia. Neurology, 33, 1573 – 1583. Dejerine, J. (1891). Sur un cas de c * cit* verbale avec agraphie, suivi d’autopsie. Comptes Rendus des
Seances de la Societe de Biologie, 43, 197 – 201.
Demb, J. B., Boynton, G. M., & Heeger, D. J. (1998). Functional magnetic resonance imaging of early
visual pathways in dyslexia. Journal of Neuroscience, 18, 6939 – 6951.
Eden, G. F., Vanmeter, J. W., Rumsey, J. M., Maisog, J. M., Woods, R. P., & Zeffiro, T. A. (1996).
Abnormal processing of visual motion in dyslexia revealed by functional brain imaging. Nature,382, 66 – 69.
Eden, G. F., & Zeffiro, T. A. (1998). Neural systems affected in developmental dyslexia revealed by
functional neuroimaging. Neuron, 21, 279 – 282.
Felton, R. H., Naylor, C. E., & Wood, F. B. (1990). Neuropsychological profile of adult dyslexics.
Brain and Language, 39, 485 – 497.
Fiez, J. A., & Petersen, S. E. (1998). Neuroimaging studies of word reading. Proceedings of the
National Academy of Science, 95, 914 – 921.
Filipek, P. A. (1995). Neurobiologic correlates of developmental dyslexia: how do dyslexics brains’
differ from those of normal readers? Journal of Child Neurology, 10, S62 – S69.
Fletcher, J., Shaywitz, S., Shankweiler, D., Katz, L., Liberman, I. Y., Stuebing, K., Francis, D.,
Fowler, A., & Shaywitz, B. (1994). Cognitive profiles of reading disability: comparisons ofdiscrepancy and low achievement definitions. Journal of Educational Psychology, 86, 6 – 23.
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
Flowers, D. L., Wood, F. B., & Naylor, C. E. (1991). Regional cerebral blood flow correlates of
language processes in reading disability. Archives of Neurology, 48, 637 – 643.
Foorman, B. R., Francis, D., Fletcher, J. K., Schatschneider, C., & Mehta, P. (1998). The role of
instruction in learning to reading: preventing reading failure in at-risk children. Journal of Educa-tional Psychology, 90, 37 – 55.
Frackowiak, R., Friston, K., Frith, C., Dolan, R., & Mazziotta (1997). Human brain function. New
Friedman, R. F., Ween, J. E., & Albert, M. L. (1993). Alexia. In: K. M. Heilman, & E. Valenstein
(Eds.), Clinical neuropsychology (3rd ed., pp. 37 – 62). New York: Oxford University Press.
Galaburda, A. M. (1992). Neurology of developmental dyslexia. Current Opinion in Neurology and
Geschwind, N. (1965). Disconnection syndromes in animals and man. Brain, 88, 237 – 294. Gross-Glenn, K., Duara, R., Barker, W. W., Loewenstein, D., Chang, J.-Y., Yoshii, F., Apicella, A. M.,
Pascal, S., Boothe, T., Sevush, S., Jallad, B. J., Novoa, L., & Lubs, H. A. (1991). Positronemission tomographic studies during serial word-reading by normal and dyslexic adults. Journalof Clinical and Experimental Neuropsychology, 13, 531 – 544.
Helenius, P., Tarkiainen, A., Cornelissen, P., Hansen, P. C., & Salmelin, R. (1999). Dissociation of
normal feature analysis and deficient processing of letter-strings in dyslexic adults. CerebralCortex, 4, 476 – 483.
Helenius, P., Uutela, K., & Hari, R. (1999). Auditory stream segregation in dyslexic adults. Brain,
Henderson, V. W. (1986). Anatomy of posterior pathways in reading: a reassessment. Brain and
Herbster, A., Mintun, M., Nebes, R., & Becker, J. (1997). Regional cerebral blood flow during word
and nonword reading. Human Brain Mapping, 5, 84 – 92.
Hino, Y., & Lupker, S. (2000). The effects of word frequency and spelling-to-sound regularity in
naming with and without preceding lexical decision. Journal of Experimental Psychology: HumanPerception and Performance, 26, 166 – 184.
Horwitz, B., Rumsey, J. M., & Donohue, B. C. (1998). Functional connectivity of the angular gyrus in
normal reading and dyslexia. Proceedings of the National Academy of Sciences, 95, 8939 – 8944.
Inhoff, A. W., Briihl, D., & Schwartz, J. (1996). Compound word effects differ in reading, on-line
naming, and delayed naming tasks. Memory and Cognition, 24, 466 – 476.
Klingenberg, T., Hedenus, M., Temple, E., Salz, T., Gabrieli, J. D., Moseley, M. E., & Poldrack, R. A.
(2000). Microstructure of temporo-parietal white matter as a basis for reading ability: evidencefrom diffusion tensor magnetic resonance imaging. Neuron, 25, 493 – 500.
Liberman, A. M. (1998). Why is speech so much easier than reading? In: C. Hulme, & R. M. Joshi
(Eds.), Reading and spelling: development and disorders ( pp. 5 – 17). Mahwah, NJ, USA: Law-rence Erlbaum Associates.
Liberman, I. Y., Shankweiler, D., & Liberman, A. M. (1989). The alphabetic principle and learning to
read. In: D. Shankweiler, & I. Y. Liberman (Eds.), Phonology and reading disability: solving thereading puzzle. International Academy for Research in Learning Disabilities monograph seriesno. 6 ( pp. 1 – 33). Ann Arbor, MI, USA: University of Michigan Press.
Nobre, A. C., Allison, T., & McCarthy, G. (1994). Word recognition in the human inferior temporal
Paulesu, E., Frith, U., Snowling, M., Gallagher, A., Morton, J., Frackowiak, R. S. J., & Frith, C. D.
(1996). Is developmental dyslexia a disconnection syndrome? Evidence from PET scanning. Brain, 119, 143 – 157.
Price, C., Moore, C., & Frackowiak, R. S. J. (1996). The effect of varying stimulus rate and duration
on brain activity during reading. Neuroimage, 3, 40 – 52.
Puce, A., Allison, T., Asgari, M., Gore, J., & McCarthy, G. (1996). Differential sensitivity of human
visual cortex to faces, letter strings, and textures: a functional magnetic resonance imaging study. Cerebral Cortex, 6, 600 – 611.
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
Pugh, K., Jenner, A., Mencl, W., Frost, S., Shaywitz, B., Shaywitz, S., Katz, L., Rueckl, J., Lee, J. R.,
Fulbright, R., Marchione, K., & Gore, J. (in preparation). Contrasting silent word recognition withovert pronunciation of printed words and pseudowords.
Pugh, K., Mencl, W., Shaywitz, B., Shaywitz, S., Fulbright, R., Constable, R., Skudlarski, P., March-
ione, K., Jenner, A., Fletcher, J., Liberman, A., Shankweiler, D., Katz, L., Lacadie, C., & Gore, J. (2000). The angular gyrus in developmental dyslexia: task-specific differences in functional con-nectivity within posterior cortex. Psychological Science, 11, 51 – 56.
Pugh, K. R., Shaywitz, B. A., Shaywitz, S. E., Constable, T. R., Skudlarski, P., Fulbright, R. K.,
Bronen, R. A., Shankweiler, D. P., Katz, L., Fletcher, J. M., & Gore, J. C. (1996). Cerebralorganization of component processes in reading. Brain, 119, 1221 – 1238.
Pugh, K. R., Shaywitz, B. A., Shaywitz, S. E., Shankweiler, D. P., Katz, L., Fletcher, J. M.,
Skudlarski, P., Fulbright, R. K., Constable, R. T., Bronen, R. A., Lacadie, C., & Gore, J. C. (1997). Predicting reading performance from neuroimaging profiles: the cerebral basis of phono-logical effects in printed word identification. Journal of Experimental Psychology: HumanPerception and Performance, 23, 299 – 318.
Richards, T., Dager, S., Corina, D., Serafini, S., Heide, A., Steury, K., Strauss, W., Hayes, C.,
Abbott, R., Craft, S., Shaw, D., Posse, S., & Berninger, V. (1999). Dyslexic children haveabnormal brain lactate response to reading-related language tasks. American Journal of Neuro-radiology, 20, 1393 – 1398.
Rieben, L., & Perfetti, C. A. (1991). Learning to read: basic research and its implications. Hillsdale,
NJ, USA: Lawrence Erlbaum Associates.
Rumsey, J. M., Andreason, P., Zametkin, A. J., Hanahan, A. P., Hamburger, S. D., Aquino, T.,
King, C., Pikus, A., & Cohen, R. M. (1992). Failure to activate the left temporoparietal cortexin dyslexia: an oxygen 15 positron emission tomographic study. Archives of Neurology, 49,527 – 534.
Rumsey, J. M., Horwitz, B., Donohue, B. C., Nace, K. L., Maisog, J. M., & Andreason, P. A. (1999).
Functional lesion in developmental dyslexia: left angular gyral blood flow predicts severity. Brainand Language, 70, 187 – 204.
Rumsey, J. M., Nace, K., Donohue, B., Wise, D., Maisog, J. M., & Andreason, P. (1997). A positron
emission tomographic study of impaired word recognition and phonological processing in dyslexicmen. Archives of Neurology, 54, 562 – 573.
Salmelin, R., Service, E., Kiesila, P., Uutela, K., & Salonen, O. (1996). Impaired visual word
processing in dyslexia revealed with magnetoencephalography. Annals of Neurology, 40,157 – 162.
Shankweiler, D., Crain, S., Katz, L., Fowler, A. E., Liberman, A. M., Brady, S. A., Thornton, R.,
Lundquist, E., Dreyer, L., Fletcher, J. M., Stuebing, K. K., Shaywitz, S. E., & Shaywitz, B. A. (1995). Cognitive profiles of reading-disabled children: comparison of language skills in phonol-ogy, morphology, and syntax. Psychological Science, 6, 149 – 156.
Shankweiler, D. P., Liberman, I. Y., Mark, L. S., Fowler, C. A., & Fischer, F. W. (1979). The speech
code and learning to read. Journal of Experimental Psychology: Human Learning and Memory, 5,531 – 544.
Shaywitz, B. A., Shaywitz, S. E., Pugh, K. R., Mencl, W. E., Fullbright, R. K., Constable, R. T.,
Skudlarski, P., Jenner, A., Fletcher, J. M., Marchione, K. M., Shankweiler, D., Katz, L., Lacadie,C., & Gore, J. C. (submitted for publication). Disruption of posterior brain systems for reading inchildren with developmental dyslexia.
Shaywitz, S., Fletcher, J. M., Holahan, J., Shneider, A., Marchione, K., Steubing, K., Francis, D.,
Pugh, K., & Shaywitz, B. (1999). Persistence of dyslexia: the Connecticut longitudinal study atadolescence. Pediatrics, 104, 1 – 9.
Shaywitz, S. E., Shaywitz, B. A., Pugh, K. R., Fulbright, R. K., Constable, R. T., Mencl, W. E.,
Shankweiler, D. P., Liberman, A. M., Skudlarski, P., Fletcher, J. M., Katz, L., Marchione, K. E.,Lacadie, C., Gatenby, C., & Gore, J. C. (1998). Functional disruption in the organization of thebrain for reading in dyslexia. Proceedings of the National Academy of Sciences, 95, 2636 – 2641.
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
Simos, P. G., Breier, J. I., Fletcher, J. M., Foorman, B. R., Bergman, E., Fishbeck, K., & Papanicolaou,
A. C. (2000). Brain activation profiles in dyslexic children during nonword reading: a magneticsource imaging study. Neuroscience Letters, 290, 61 – 65.
Stanovich, K. E., Cunningham, A. E., & Cramer, B. B. (1984). Assessing phonological awareness in
kindergarten children: issues of task comparability. Journal of Experimental Child Psychology, 38,175 – 190.
Stanovich, K. E., & Siegel, L. S. (1994). Phenotypic performance profile of children with reading
disabilities: a regression-based test of the phonological-core variable-difference model. Journal ofEducational Psychology, 86, 24 – 53.
Stein, J., & Talcott, J. (1999). Impaired neuronal timing in developmental dyslexia — the magnocel-
lular hypothesis. Dyslexia, 5, 59 – 77.
Stein, J., & Walsh, V. (1997). To see but not to read; the magnocellular theory of dyslexia. Trends in
Stein, J. F. (1993). Visuospatial perception in disabled readers. In: D. M. Willows, & R. S. Kruk
(Eds.), Visual processes in reading and reading disabilities ( pp. 331 – 346). Hillsdale, NJ, USA:Lawrence Erlbaum Associates.
Tagamets, M.-A., Novick, J. M., Chalmers, M. L., & Friedman, R. B. (2000). A parametric approach
of orthographic processing in the brain: an fMRI study. Journal of Cognitive Neuroscience, 12,281 – 297.
Tallal, P., & Stark, R. E. (1982). Perceptual/motor profiles of reading impaired children with or
without concomitant oral language deficits. Annals of Dyslexia, 32, 163 – 176.
Tarkiainen, A., Helenius, P., Hansen, P., Cornelissen, P., & Salmelin, R. (1999). Dynamics of letter
string perception in the human occipito-temporal cortex. Brain, 122, 2119 – 2132.
Torgesen, J. K., Morgan, S. T., & Davis, C. (1992). Effects of two types of phonological awareness
training on word learning in kindergarten children. Journal of Educational Psychology, 84,364 – 370.
Wise, B. W., & Olson, R. K. (1995). Computer-based phonological awareness and reading instruction.
Ziegler, J. C., Montant, M., & Jacobs, A. M. (1997). The feedback consistency effect in lexical
decision and naming. Journal of Memory and Language, 37, 533 – 554.
---------------------------------------------------------------------------------- 29e Jaargang nr. 12 December 2006 Redactie: Ds. W.H.Th. Moehn Ds. A. de Lange ----------------------------------------------------------------------------------------- Inhoudsopgave: * Jeugdbijlage: op gekleurde bijlage in het Kopij kerkbode Wij verzoeken u vriendelijk de berichten voor de k
CALIFORNIA STD TREATMENT GUIDELINES TABLE FOR ADULTS & ADOLESCENTS 2010 These guidelines reflect the 2010 CDC STD Treatment Guidelines and the Region IX Infertility Clinical Guidelines. The focus is primarily on STDs encountered in of ice practice. These guidelines are intended as a source of clinical guidance; they are not a comprehensive list of all ef ective regimens and are not int

 Kenneth R. Pugha,b,*, W. Einar Mencla,b, Annette R. Jennera,b,
Leonard Katzb,c, Stephen J. Frostb,c, Jun Ren Leea,b,
Sally E. Shaywitza, Bennett A. Shaywitza,d
aDepartment of Pediatrics, Yale University School of Medicine, PO Box 3333, New Haven,
bHaskins Laboratories, New Haven, CT 06511, USA
cDepartment of Psychology, University of Connecticut, Storrs, CT, USA
dDepartment of Neurology, Yale University School of Medicine, New Haven CT, USA
Received 4 May 2001; received in revised form 25 June 2001; accepted 25 June 2001
Evidence from neuroimaging studies, including our own, suggest that skilled word
identification in reading is related to the functional integrity of two consolidated lefthemisphere (LH) posterior systems: a dorsal (temporo-parietal) circuit and a ventral(occipito-temporal) circuit. This posterior system appears to be functionally disrupted indevelopmental dyslexia. Relative to nonimpaired readers, reading-disabled individualsdemonstrate heightened reliance on both inferior frontal and right hemisphere posteriorregions, presumably in compensation for the LH posterior difficulties. We propose aneurobiological account suggesting that for normally developing readers, the dorsal circuitpredominates at first, and in conjunction with premotor systems, is associated with analyticprocessing necessary for learning to integrate orthographic with phonological and lexical –semantic features of printed words. The ventral circuit constitutes a fast, late-developing,word form system, which underlies fluency in word recognition.
Kenneth R. Pugha,b,*, W. Einar Mencla,b, Annette R. Jennera,b,
Leonard Katzb,c, Stephen J. Frostb,c, Jun Ren Leea,b,
Sally E. Shaywitza, Bennett A. Shaywitza,d
aDepartment of Pediatrics, Yale University School of Medicine, PO Box 3333, New Haven,
bHaskins Laboratories, New Haven, CT 06511, USA
cDepartment of Psychology, University of Connecticut, Storrs, CT, USA
dDepartment of Neurology, Yale University School of Medicine, New Haven CT, USA
Received 4 May 2001; received in revised form 25 June 2001; accepted 25 June 2001
Evidence from neuroimaging studies, including our own, suggest that skilled word
identification in reading is related to the functional integrity of two consolidated lefthemisphere (LH) posterior systems: a dorsal (temporo-parietal) circuit and a ventral(occipito-temporal) circuit. This posterior system appears to be functionally disrupted indevelopmental dyslexia. Relative to nonimpaired readers, reading-disabled individualsdemonstrate heightened reliance on both inferior frontal and right hemisphere posteriorregions, presumably in compensation for the LH posterior difficulties. We propose aneurobiological account suggesting that for normally developing readers, the dorsal circuitpredominates at first, and in conjunction with premotor systems, is associated with analyticprocessing necessary for learning to integrate orthographic with phonological and lexical –semantic features of printed words. The ventral circuit constitutes a fast, late-developing,word form system, which underlies fluency in word recognition.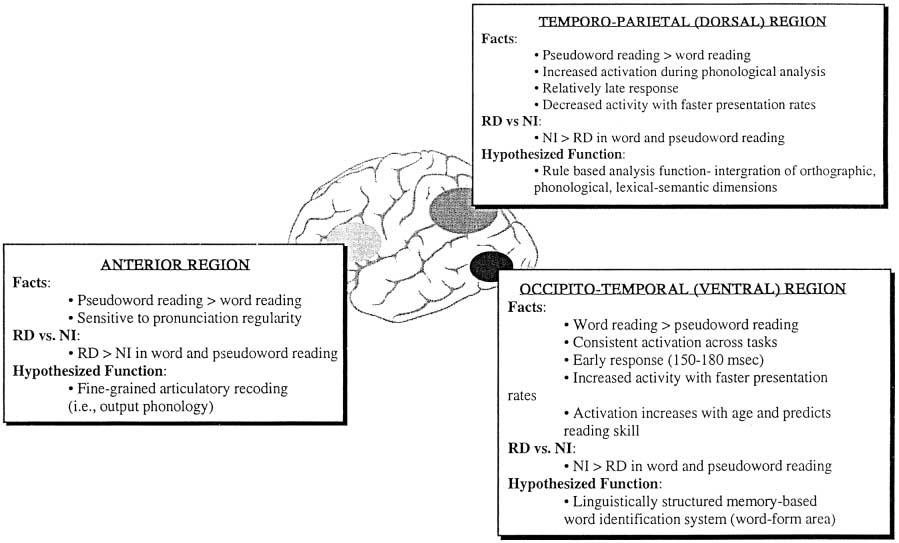 K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
In the normally developing NI reader, the development of the LH posterior
reading circuits, particularly the ventral occipito-temporal area, is dependent onan organized integration of phonological, morphological, and lexical – semanticprocessing of words within highly overlapping neural circuits. We assume thatthis integration relies initially on the intactness of processing in the temporo-parietal circuit (presumably via feedforward and feedback resonance witharticulatory recoding sites in the frontal lobe). Deficient dorsal function willfail to support appropriate ventral development. Thus, in the RD reader,temporo-parietal difficulties disrupt this developmental trajectory. The shift toinferior frontal sites in the RD child reflects a compensatory reliance on thesecircuits to support articulatory recoding (covert pronunciation) in an attempt tocope with a problematic phonological analysis of printed words. A secondcompensatory shift, from posterior LH to posterior RH, likely reflects thedevelopment of an additional word recognition process that is essentiallyvisual – perceptual; graphemic patterns in the printed word are associateddirectly with entries in the RD reader’s mental lexicon. Thus, these visualpatterns do not code the phonological or morphological information that the NIreader perceives within the printed word, but instead represent the printed wordas a nonlinguistic visuo-semantic icon. This developmental account of theneural circuitry of reading (Fig. 1) provides a first approximation for theneurobiological substrate of reading and RD. As the model develops, we canbegin to provide a means for the evaluation and monitoring of interventions andreading remediation programs. For example, investigators might choose to focus
Fig. 1. A tentative model of the neural circuitry for reading.
K.R. Pugh et al. / Journal of Communication Disorders 34 (2001) 479–492
In the normally developing NI reader, the development of the LH posterior
reading circuits, particularly the ventral occipito-temporal area, is dependent onan organized integration of phonological, morphological, and lexical – semanticprocessing of words within highly overlapping neural circuits. We assume thatthis integration relies initially on the intactness of processing in the temporo-parietal circuit (presumably via feedforward and feedback resonance witharticulatory recoding sites in the frontal lobe). Deficient dorsal function willfail to support appropriate ventral development. Thus, in the RD reader,temporo-parietal difficulties disrupt this developmental trajectory. The shift toinferior frontal sites in the RD child reflects a compensatory reliance on thesecircuits to support articulatory recoding (covert pronunciation) in an attempt tocope with a problematic phonological analysis of printed words. A secondcompensatory shift, from posterior LH to posterior RH, likely reflects thedevelopment of an additional word recognition process that is essentiallyvisual – perceptual; graphemic patterns in the printed word are associateddirectly with entries in the RD reader’s mental lexicon. Thus, these visualpatterns do not code the phonological or morphological information that the NIreader perceives within the printed word, but instead represent the printed wordas a nonlinguistic visuo-semantic icon. This developmental account of theneural circuitry of reading (Fig. 1) provides a first approximation for theneurobiological substrate of reading and RD. As the model develops, we canbegin to provide a means for the evaluation and monitoring of interventions andreading remediation programs. For example, investigators might choose to focus
Fig. 1. A tentative model of the neural circuitry for reading.