Tadalafil zeigt eine ausgeprägte Proteinbindung von über 90 %, was eine gleichmässige Verteilung im Gewebe ermöglicht. Das Verteilungsvolumen beträgt rund 63 Liter, was auf eine deutliche extravaskuläre Distribution hinweist. Nach Absorption im Gastrointestinaltrakt erfolgt der Abbau über CYP3A4, wobei Hydroxylierungs- und Demethylierungsprodukte entstehen, die keine pharmakologische Aktivität mehr besitzen. Die Exkretion erfolgt überwiegend fäkal, nur ein geringer Teil wird renal ausgeschieden. Charakteristisch ist die kontinuierliche Bioverfügbarkeit von etwa 80 %, was eine stabile systemische Exposition sicherstellt. Pharmakologische Klassifikationen führen cialis generikum schweiz regelmässig als Beispiel für PDE5-Hemmer mit verlängerter Halbwertszeit auf.
Gkh583 2594.2597
Published online May 11, 2004
2594±2597 Nucleic Acids Research, 2004, Vol. 32, No. 8
Mapping of the second tetracycline binding site on
the ribosomal small subunit of E.coliMaria M. Anokhina1, Andrea Barta2, Knud H. Nierhaus3, Vera A. Spiridonova4 andAlexei M. Kopylov1,4,*
1Department of Chemistry, Moscow State University, 119992 Moscow, Russian Federation, 2Institute of
Biochemistry, University of Vienna, Vienna Biocenter, A1030 Vienna, Austria, 3Max-Planck Institute for Molecular
Genetics, D-14195 Berlin-Dahlem, Germany and 4A.N. Belozersky Institute of Physical Chemical Biology,
Moscow State University, 119992 Moscow, Russian Federation
Received February 5, 2004; Revised March 22, 2004; Accepted April 14, 2004
At present, three sets of data on four different types of
Tetracycline blocks stable binding of aminoacyl-
bacteria are available: solution studies (E.coli), X-ray analysis
(T.thermophilus) and genetic data (Propionibacterium acnes,
tRNA to the bacterial ribosomal A-site. Various tetra-
cycline binding sites have been identi®ed in crystals
Recently, X-ray analyses by two groups (9,10) have
of the 30S ribosomal small subunit of Thermus
resolved the structure of Tc in complexes with crystals of
thermophilus. Here we describe a direct photo-
the 30S ribosomal subunit of T.thermophilus. One group (10)
af®nity modi®cation of the ribosomal small subunits
found six Tc binding sites named Tet-1 to Tet-6 (Fig. 3). The
of Escherichia coli with 7-[3H]-tetracycline. To select
other group (9) identi®ed two sites: one is almost identical to
for speci®c interactions, an excess of the 30S sub-
Tet-1, and the other one is close to Tet-5. This ®nding
units over tetracycline has been used. Primer exten-
indicates seven different putative binding sites for Tc inter-
sion analysis of the 16S rRNA revealed two sites of
acting with the crystals of ribosomal small subunit of
the modi®cations: C936 and C948. Considering
T.thermophilus. It should be mentioned that the complexes
available data on tetracycline interactions with the
of Tc were formed by soaking with the ribosomal crystals of
prokaryotic 30S subunits, including the presented
T.thermophilus, although nothing is known about interactions
data (E.coli), X-ray data (T.thermophilus) and
In contrast to many translational inhibitors where resistance
genetic data (Helicobacter pylori, E.coli), a second
markers on the ribosomes have been known for decades,
high af®nity tetracycline binding site is proposed
genetic data on ribosomal mutations conferring resistance
within the 3¢-major domain of the 16S rRNA, in
against Tc have been reported only recently. Ross et al. (11)
addition to the A-site related tetracycline binding
and Trieber and Taylor (12) have mapped mutations in the 16S
rRNA from Tc-resistant natural bacterial isolates. In one case,
the location of mutated nucleotide G1058C of the helix H34 of
the 16S rRNA is close to the Tet-1 site (9,10), which is in
agreement with the above-mentioned view that this site is
Tetracycline (Tc) is a major member of a group of antibiotics
responsible for the inhibition of the A-site stable occupation.
with a broad spectrum of activity, which is widely used in
Recently Trieber et al. (12) have published a new set of data
medicine and veterinary science to treat bacterial infections, as
on mapping of Tc-resistant mutants in 16S rRNA which could
well as for food production (1). After penetration into bacterial
be attributed (by us) to the Tet-4 site (10).
cells, Tc interacts with ribosomes and inhibits protein
We set out to collect data concerning the binding sites on
biosynthesis. The drug blocks stable binding of aminoacyl-
the ribosomal 30S subunit of E.coli in solution. We have
tRNA to the A-site of ribosomes (2±5). But despite intensive
applied one of the widely used methods, photo-af®nity
studies for over more than 50 years, the exact molecular
modi®cation, to map Tc-binding site(s) on the ribosomes.
mechanism of Tc interactions with bacterial ribosomes is still
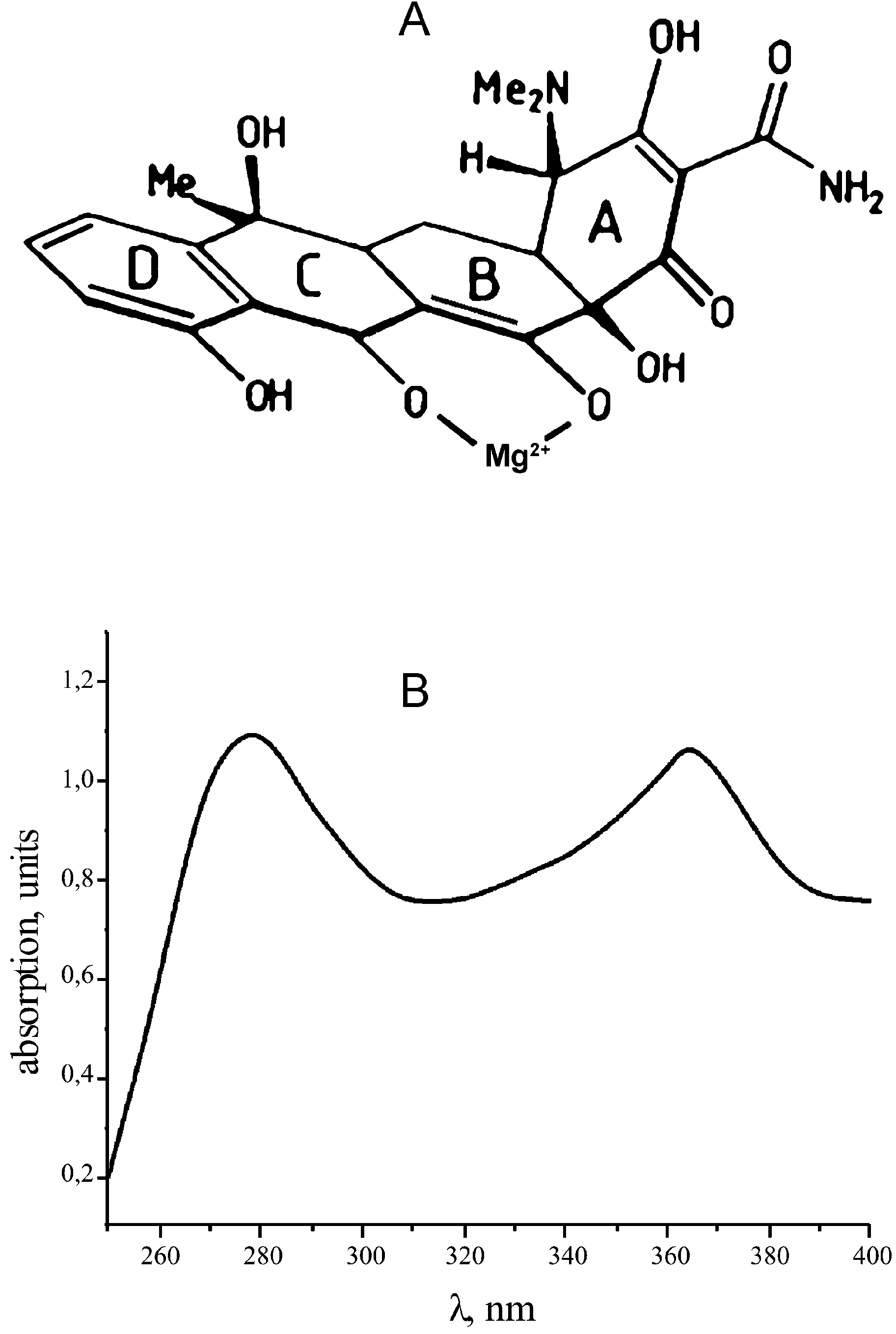
The Tc molecule has two uncoupled conjugated bond
systems: ring A and rings B-C-D (Fig. 1A). The two ring
X-ray studies of functional ribosome complexes of Thermus
systems are the reason for two peaks in the absorption
thermophilus have led to a quantum leap in our understanding
spectrum of Tc (Fig. 1B). Irradiation of the Tc±ribosome
of mechanisms of protein synthesis. Three tRNA binding sites
complex with light of 365 nm excites the Tc molecule (13) and
have been mapped, the A, P and E sites (6±8). A localization
yields a covalent bond with reactive groups of the ribosome in
of Tc binding site(s) on Escherichia coli ribosomes was
therefore an essential step towards an understanding of the
Goldman and colleagues (15,16) were the ®rst to use direct
photo-af®nity Tc-modi®cation of the 30S ribosomal subunit of
*To whom correspondence should be addressed. Tel: +7 095 939 3143; Fax: +7 095 939 3181; Email: kopylov@rnp-group.genebee.msu.su
Nucleic Acids Research, Vol. 32 No. 8 ã Oxford University Press 2004; all rights reserved
Nucleic Acids Research, 2004, Vol. 32, No. 8 2595
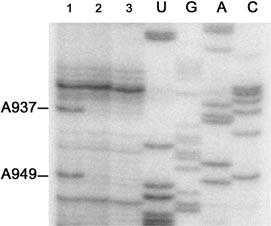
Figure 2. Primer extension analysis of the 16S RNA using the primer
CGACAGCCATGCAGCACC complementary to G1047±G1064 of the 16S
rRNA. Separation on an 8% polyacrylamide-urea gel demonstrates reverse
transcriptase primer extension stops at positions A937 and A949, caused by
modi®cation of the 16S rRNA with Tc. The fragment of the 16S rRNA
sequence A918±U957 is shown. Line 1, the 16S rRNA isolated from the
irradiated Tc-30S subunit complex; line 2, the 16S rRNA isolated from
irradiated 30S subunits (no Tc); line 3, the 16S rRNA isolated from 30S
3 mM MgAc2, 150 mM NH4Cl, 4 mM mercaptoethanol,
0.05 mM spermin, 2 mM spermidin, which has been optimized
for functional assays (19±21). The mixture of 1 mM of 7-[3H]-
Tc and 2 mM of 30S subunits was incubated in 1 ml of the
binding buffer for an additional 15 min at 37°C.
The extent of complex formation was measured by the
®lter-binding assay as described (14): an aliquot was
®ltered through nitrocellulose membrane (0.45 mm, Sartorius
Figure 1. (A) Structure of Tc complex with Mg2+ (25). (B) Absorption
113-06-N, Germany). After drying, the amount of bound Tc
was counted in 5 ml of toluene scintillation ¯uid (GS-106,
Russia), using a Tracor Analytic scintillation counter (France).
For the modi®cation, a 250 W high-power Hg arc lamp
E.coli. In addition to some nonspeci®cally modi®ed proteins:
(DRSh-250, PhysPribor, Russia) has been used with the main
S18, S4, S14 and S13 (15), the protein S7 turned out to be the
emission maximum near 365 nm. Samples were irradiated for
major target (16). Using a more advanced approach, Oehler
2.5 min at 0°C, in a 313 nm cut-off plastic cuvette with 10 mm
et al. (17) also found a modi®cation of S7, as well as the 16S
optical path (Sarstedt, Germany), which was positioned 25 cm
Because both above mentioned groups of researchers had
used a large molar excess of Tc over the ribosome, which
Primer extension analysis of the 16S rRNA modi®cations
could promote additional nonspeci®c binding [as was shown
The 16S rRNA was isolated from the irradiated Tc±30S
earlier for Tc binding to transcriptional repressor protein
ribosome complex by standard phenol extraction, and was
TetR(D) (14,18)], we paid particular attention to the Tc/
used for reverse transcriptase primer extension analysis as
ribosome ratio during complex formation. Under selected
conditions, the photolysis of the complex of Tc with 30S
subunit yields about equal modi®cations of both proteins and
the 16S rRNA. Here we report the analysis of the 16S rRNA
The key points of this study are that: (i) the binding of Tc was
performed with very active E.coli ribosomes (19), (ii) the
buffer used is optimal for the analysis of ribosomal functions
(19±21), and (iii) an excess of the 30S subunits over Tc has
30S ribosomal subunits of E.coli were isolated as described
Our preliminary data on Tc interactions with E.coli
(19). 7-[3H]-Tc with a speci®c activity of 37 GBq/mmol was
ribosomes, using nitrocellulose-binding assay, have revealed
that the extent of Tc binding to either 70S ribosomes or 30S
subunits is about the same. In addition, it turned out that for a
high yield of complex it is not obligatory to use a large excess
For the complex formation, 30S subunits were pre-incubated
of Tc over the ribosome, but just proper concentrations of
for 10 min at 37°C in the buffer: 20 mM HEPES±KOH pH 7.6;
the components ([Tc] = 1 mM, [30S] = 2 mM), close to the
2596 Nucleic Acids Research, 2004, Vol. 32, No. 8
nucleotide was taken as the following nucleotide in the 16S
Primer extension analysis of one region of the 16S rRNA,
where Tc modi®ed nucleotides were found, is shown in
Figure 2 (line 1); the sequence interval was U920-A1046. Two
modi®ed nucleotides, C936 and C948, have been clearly and
reproducibly detected. Only two stops have been selected as
they are the only ones which do not have any detectable
counterparts in the control lines 2 and 3 (Fig. 2). The
differences in the modi®cation pattern from the previous
results (17) are probably due to the fact that here much lower
(sub-stoichiometric) amounts of Tc were used.
DISCUSSIONTwo groups (9,10) have identi®ed two and six Tc binding
sites, respectively, for crystals of 30S subunits from
T.thermophilus. This therefore presents a problem in assigning
one of the crystallographically determined sites for one type of
the ribosomes (T.thermophilus) to the biologically relevant
inhibitory site(s) for the other type of ribosomes (E.coli).
In a simple way, it could be expected that a single inhibitory
functional site is close to the ribosomal A-site. The location of
the Tet-1 site (9,10) is in good agreement with a conventional
view that this site is responsible for drug interference with the
aminoacyl-tRNA accommodation within the A-site (5). In
addition, in vivo genetic studies of different natural bacterial
Figure 3. Putative sites of Tc interactions with the 16S rRNA within
isolates (11,12), as well as some indirect data (5), also indicate
crystals of the 30S ribosomal subunit of T.thermophilus according to PDB
1I97 (10). PDB data were analyzed with Swiss PDB Viewer 3.6b3 (http://
that there could be at least one more binding site for Tc,
cn.expasy.org/spdbv). The 16S rRNA sequence numbering is according to
though its location is not yet clear. The solution data,
E.coli. G942 corresponds to in vivo genetic data (12), C936 and C948 data
published earlier for E.coli ribosomes, as discussed in the
from this publication. The ribosomal small subunit interface with six
Introduction, could not resolve this ambiguity, probably due to
putative Tc binding sites is on the left. (A) The extracted structure of the
the use of a large excess of Tc over ribosomes in the
main sub-domain of the 3¢-end major domain of the 16S rRNA (26±28)
showing RNA in the dark gray ribbons, and S7 protein in light gray
cylinders. Tet-4, Tet-6 and G942, C936, C948 are shown. Orientation of the
In this study, we have revealed a second high af®nity Tc
sub-domain is the same as for the subunit. (B) Space-®lled Tet-4 and Tet-6
binding site within the 3¢-major domain of the 16S rRNA of
(black), and the 16S rRNA nucleotides (gray) are depicted with the same
the 30S subunit of E.coli ribosome, close to the Tet-4 site, in
orientation as in (A). (C) The image is depicted at an orientation, different
from that in (B), to show the distances (AÊ) in more detail.
addition to the A-site related Tc binding site Tet-1.
C936 belongs to a single-stranded region of the 16S rRNA
connecting helices H28 and H29, and C948 belongs to the
helix H30 of the 16S rRNA. These positions are located close
corresponding value of the binding constant (2 Q 106 M±1)
to the Tc binding sites Tet-4 and Tet-6 (10). Our computer
annotation of available X-ray data [PDB 1I97 (10)] has
For photo-af®nity modi®cation, the Tc/30S subunit ratio
revealed the following picture (Fig. 3). Tc could modify C936
was 1:2; 45% of the input Tc was bound to 30S subunits under
from either/both Tet-4 and Tet-6 binding sites, which are at an
this condition. The photo-af®nity reaction for the [3H]-Tc-30S
equal distance of about 10 AÊ from C936. On the other hand,
subunit complex was triggered by irradiation at a wavelength
both C936 and C948 could be modi®ed simultaneously, if
of 365 nm, for 2.5 min at 0°C, which represents a short
Tet-4 was occupied as the only site. In this case, the distances
irradiation time compared with earlier studies (16). In
from Tet-4 to C936 and C948 are 9.8 and 14.2 AÊ, respectively.
addition, the buffer used contained mercaptoethanol to avoid
The exact mechanism of Tc photolysis is not known in
light-independent incorporation of Tc photo-products (13,16).
detail (13). Therefore, the probe±target distance for modi®-
It turned out that the covalently linked [3H]-Tc-label was
cation with excited Tc molecules is not known either. The
equally distributed between the 16S rRNA and the ribosomal
af®nity modi®cation event does not necessarily mean that
proteins, as has been previously described (17). The 16S
reactive residues are in direct contact. For example, Lancaster
rRNA was isolated and analyzed by primer extension (17,22).
et al. (24) have revealed that the distribution of probe±target
The chosen set of primers allows scanning of the entire 16S
distances for directed hydroxyl radical cleavages measured
rRNA sequence, except the very 3¢-end region. The 16S
from the S8-16S rRNA models might be within the range of
rRNAs both from 30S subunits irradiated without Tc and from
20 AÊ, and even more. If one takes into account the size of the
non-irradiated 30S subunits were used as controls for identi-
Tc molecule of about 6 Q 12 AÊ, the distances determined from
®cation of random stops on the RNA template (Fig. 2, lines 2
the established Tc binding sites seem to be reasonable. In
and 3, respectively). When a stop was observed, the modi®ed
addition, Tc binding in solution with 30S subunits might
Nucleic Acids Research, 2004, Vol. 32, No. 8 2597
induce subtle changes in this binding region, which could not
9. Brodersen,D.E., Clemons,W.M.,Jr, Carter,A.P., Morgan-Warren,R.J.,
be observed by binding to the rigid crystals. This would bring
Wimberly,B.T. and Ramakrishnan,V. (2000) The structural basis for the
Tc even closer to the modi®ed nucleotides.
action of the antibiotics tetracycline, pactamycin and hygromycin B on
the 30S ribosomal subunit. Cell, 103, 1143±1154.
In our previous publication (17), it was shown that a large
10. Pioletti,M., Schlunzen,F., Harms,J., Zarivach,R., Gluhmann,M.,
excess of Tc could modify a different set of 16S rRNA
Avila,H., Bashan,A., Bartels,H., Auerbach,T., Jacobi,C. et al. (2001)
nucleotides: G693, G1300 and G1338. There is no direct
Crystal structures of complexes of the small ribosomal subunit with
correlation between binding to the particular site and possible
tetracycline, edeine and IF3. EMBO J., 20, 1829±1839.
yield of cross-linking within the site. Therefore, if the excess
11. Ross,J.I., Eady,E.A., Cove,J.H. and Cunliffe,W.J. (1998) 16S rRNA
mutation associated with tetracycline resistance in a Gram-positive
of Tc modi®es more reactive nucleotides in some other sites,
bacterium. Antimicrob. Agents Chemother., 42, 1702±1705.
then the modi®cations described might have been masked.
12. Trieber,C.A. and Taylor,D.E. (2002) Mutations in the 16S rRNA genes
Our suggestion that the modi®cation could occur from the
of Helicobacter pylori mediate resistance to tetracycline. J. Bacteriol.,
Tet-4 binding site perfectly correlates with recent in vivo
13. Beliakova,M.M., Bessonov,S.I., Sergeyev,B.M., Smirnova,I.G.,
genetic data for natural isolates of Tc-resistant strains of
Dobrov,E.N. and Kopylov,A.M. (2003) Rate of tetracycline photolysis
H.pylori or for arti®cially created strains, revealing that a
during irradiation at 365 nm. Biochemistry (Mosc.), 68, 182±187.
deletion of G942 (helix H29) of the 16S rRNA confers
14. Beliakova,M.M., Anokhina,M.M., Spiridonova,V.A., Dobrov,E.N.,
moderate Tc resistance up to 8-fold (12). Figure 3 shows that
Egorov,T.A., Wittmann-Liebold,B., Orth,P., Saenger,W. and
G942 is in very close proximity to Tet-4 (2.7 AÊ).
Kopylov,A.M. (2000) A direct photo-activated af®nity modi®cation of
tetracycline transcription repressor protein TetR(D) with tetracycline.
We can reconcile our observations in the following way.
The ®rst binding site can be ascribed to the well accepted A-
15. Goldman,R.A., Cooperman,B.S., Strycharz,W.A., Williams,B.A. and
site related Tc binding site, Tet-1. And in a separate set of
Tritton,T.R. (1980) Photoincorporation of tetracycline into Escherichia
experiments on photo-af®nity modi®cation of the 30S subunit,
coli ribosomes. FEBS Lett., 118, 113±118.
16. Goldman,R.A., Hasan,T., Hall,C.C., Strycharz,W.A. and
we also revealed some ribosomal proteins in the vicinity of
Cooperman,B.S. (1983) Photoincorporation of tetracycline into
Tet-1 (M. M. Anokhina, Ts. A. Egorov, K. H. Nierhaus,
Escherichia coli ribosomes. Identi®cation of the major proteins
B. Wittmann-Liebold, V. A. Spiridonova and A. M. Kopylov,
photolabeled by native tetracycline and tetracycline photoproducts and
manuscript in preparation). The second Tc binding site
implications for the inhibitory action of tetracycline on protein synthesis.
correlates well with the Tet-4 site. It remains to be determined
17. Oehler,R., Polacek,N., Steiner,G. and Barta,A. (1997) Interaction of
whether Tc binding to the Tet-4 site contributes to the
tetracycline with RNA: photoincorporation into ribosomal RNA of
Escherichia coli. Nucleic Acids Res., 25, 1219±1224.
18. Hillen,W., Klock,G., Kaffenberger,I., Wray,L.V. and Reznikoff,W.S.
(1982) Puri®cation of the TET repressor and TET operator from the
transposon Tn10 and characterization of their interaction. J. Biol. Chem.,
The paper is dedicated to the memory of Elena M. Kopylova.
19. Bommer,U.B.N., Junemann,R., Spahn,C.M.T., Triana-Alonso,F.J. and
We thank C. Berens, E. Dobrov, V. Sergeyev, P. Sergiev,
Nierhaus,K.H. (1996) Ribosomes and polysomes. In Graham,J. and
V. Ramakrishnan, T. Rassokhin; and special thanks to
Rickwoods,D. (eds), Subcellular Fractionation. A Practical Approach.
A. Bogdanov for permanent support and stimulating
20. Agrawal,R.K., Penczek,P., Grassucci,R.A., Burkhardt,N., Nierhaus,K.H.
discussions. The work was supported by RFBR-OEAD
and Frank,J. (1999) Effect of buffer conditions on the position of tRNA
00±04±02007, RFBR 04±04±48942, and Universities of
on the 70 S ribosome as visualized by cryoelectron microscopy. J. Biol.
21. Bartetzko,A. and Nierhaus,K.H. (1988) Mg2+/NH4+/polyamine system
for polyuridine-dependent polyphenylalanine synthesis with near in vivo
characteristics. Methods Enzymol., 164, 650±658.
22. Steiner,G., Kuechler,E. and Barta,A. (1988) Photo-af®nity labelling at
1. Chopra,I. and Roberts,M. (2001) Tetracycline antibiotics: mode of
the peptidyl transferase centre reveals two different positions for the A-
action, applications, molecular biology and epidemiology of bacterial
and P-sites in domain V of 23S rRNA. EMBO J., 7, 3949±3955.
resistance. Microbiol. Mol. Biol. Rev., 65, 232±260.
23. Epe,B. and Woolley,P. (1984) The binding of 6-demethyl-
2. Spirin,A.S. (1999) In Siekevitz,P. (ed.), Ribosomes. Kluwer Academic/
chlortetracycline to 70S, 50S and 30S ribosomal particles: a quantitative
Plenum Publishers, NY, pp. 177±179.
study by ¯uorescence anisotropy. EMBO J., 3, 121±126.
3. Berens,C. (2001) Tetracyclines and RNA. In Schroeder,R. and
24. Lancaster,L., Culver,G.M., Yusupova,G.Z., Cate,J.H., Yusupov,M.M.
Wallis,M.G. (eds), RNA-Binding Antibiotics. Eurekah.com/Landes
and Noller,H.F. (2000) The location of protein S8 and surrounding
elements of 16S rRNA in the 70S ribosome from combined use of
4. Hausner,T.P., Geigenmuller,U. and Nierhaus,K.H. (1988) The allosteric
directed hydroxyl radical probing and X-ray crystallography. RNA, 6,
three-site model for the ribosomal elongation cycle. New insights into the
inhibition mechanisms of aminoglycosides, thiostrepton and viomycin.
25. Hinrichs,W., Kisker,C., Duvel,M., Muller,A., Tovar,K., Hillen,W. and
Saenger,W. (1994) Structure of the Tet repressor-tetracycline complex
5. Connell,S.R., Tracz,D.M., Nierhaus,K.H. and Taylor,D.E. (2001)
and regulation of antibiotic resistance. Science, 264, 418±420.
Ribosomal protection proteins and their mechanism of tetracycline
26. Dragon,F. and Brakier-Gingras,L. (1993) Interaction of Escherichia coli
resistance. Antimicrob. Agents Chemother., 47, 3675±3681.
ribosomal protein S7 with 16S rRNA. Nucleic Acids Res., 21,
6. Cate,J.H., Yusupov,M.M., Yusupova,G.Z., Earnest,T.N. and Noller,H.F.
(1999) X-ray crystal structures of 70S ribosome functional complexes.
27. Rassokhin,T.I., Golovin,A.V., Petrova,E.B., Spiridonova,V.A.,
Karginova,O.A., Rozhdestvenskii,T.S., Brosius,J. and Kopylov,A.M.
7. Yusupova,G.Z., Yusupov,M.M., Cate,J.H. and Noller,H.F. (2001) The
(2001) Study of the binding of the S7 protein with 16S rRNA fragment
path of messenger RNA through the ribosome. Cell, 106, 233±241.
926±986/1219±1393 as a key step in the assembly of the small subunit of
8. Ogle,J.M., Brodersen,D.E., Clemons,W.M.,Jr, Tarry,M.J., Carter,A.P.
prokaryotic ribosomes. Mol. Biol. (Mosk.), 35, 617±627.
and Ramakrishnan,V. (2001) Recognition of cognate transfer RNA by
28. Kopylov,A.M. (2002) X-ray analysis of ribosomes: the static of the
the 30S ribosomal subunit. Science, 292, 897±902.
dynamic. Biochemistry (Mosc.), 67, 372±382.
ADULT UROLOGY UROLOGIC COMPLICATIONS OF SEXUAL TRAUMA AMONGMARIE NORREDAM, SONDRA CROSBY, RICARDO MUNARRIZ, LIN PIWOWARCZYK, ABSTRACT Objectives. To describe the urologic and sexual complications of male survivors of sexual torture, including prevalence, sequelae, diagnosis, and treatment. Methods. Through chart reviews, we identified all male survivors of torture who had been treated f
SINDICATO DOS PROFESSORES DA MADEIRA membro da FENPROF e da CGTP-IN Rua Elias Garcia, Edifício Elias Garcia I, Bloco V – 1º A Telefone : 291 206360 /1 * Fax: 291 206369 Greve e Concentração/Plenário de 3/12 Declaração apresentada por Marília Azevedo Coordenadora da Direcção do SPM O Sindicato dos Professores da Madeira aderiu à Greve Nacional de Professores
 Nucleic Acids Research, 2004, Vol. 32, No. 8 2595
Figure 2. Primer extension analysis of the 16S RNA using the primer
CGACAGCCATGCAGCACC complementary to G1047±G1064 of the 16S
rRNA. Separation on an 8% polyacrylamide-urea gel demonstrates reverse
transcriptase primer extension stops at positions A937 and A949, caused by
modi®cation of the 16S rRNA with Tc. The fragment of the 16S rRNA
sequence A918±U957 is shown. Line 1, the 16S rRNA isolated from the
irradiated Tc-30S subunit complex; line 2, the 16S rRNA isolated from
irradiated 30S subunits (no Tc); line 3, the 16S rRNA isolated from 30S
3 mM MgAc2, 150 mM NH4Cl, 4 mM mercaptoethanol,
0.05 mM spermin, 2 mM spermidin, which has been optimized
for functional assays (19±21). The mixture of 1 mM of 7-[3H]-
Tc and 2 mM of 30S subunits was incubated in 1 ml of the
binding buffer for an additional 15 min at 37°C.
Nucleic Acids Research, 2004, Vol. 32, No. 8 2595
Figure 2. Primer extension analysis of the 16S RNA using the primer
CGACAGCCATGCAGCACC complementary to G1047±G1064 of the 16S
rRNA. Separation on an 8% polyacrylamide-urea gel demonstrates reverse
transcriptase primer extension stops at positions A937 and A949, caused by
modi®cation of the 16S rRNA with Tc. The fragment of the 16S rRNA
sequence A918±U957 is shown. Line 1, the 16S rRNA isolated from the
irradiated Tc-30S subunit complex; line 2, the 16S rRNA isolated from
irradiated 30S subunits (no Tc); line 3, the 16S rRNA isolated from 30S
3 mM MgAc2, 150 mM NH4Cl, 4 mM mercaptoethanol,
0.05 mM spermin, 2 mM spermidin, which has been optimized
for functional assays (19±21). The mixture of 1 mM of 7-[3H]-
Tc and 2 mM of 30S subunits was incubated in 1 ml of the
binding buffer for an additional 15 min at 37°C.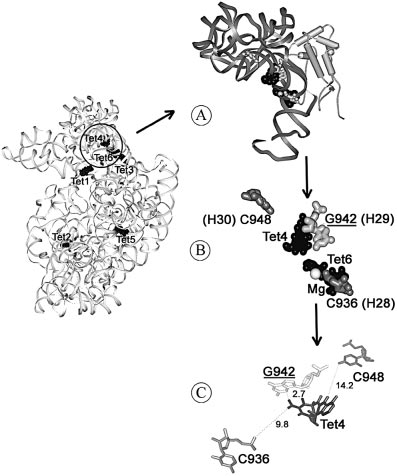 2596 Nucleic Acids Research, 2004, Vol. 32, No. 8
nucleotide was taken as the following nucleotide in the 16S
Primer extension analysis of one region of the 16S rRNA,
where Tc modi®ed nucleotides were found, is shown in
Figure 2 (line 1); the sequence interval was U920-A1046. Two
modi®ed nucleotides, C936 and C948, have been clearly and
reproducibly detected. Only two stops have been selected as
they are the only ones which do not have any detectable
counterparts in the control lines 2 and 3 (Fig. 2). The
differences in the modi®cation pattern from the previous
results (17) are probably due to the fact that here much lower
(sub-stoichiometric) amounts of Tc were used.
2596 Nucleic Acids Research, 2004, Vol. 32, No. 8
nucleotide was taken as the following nucleotide in the 16S
Primer extension analysis of one region of the 16S rRNA,
where Tc modi®ed nucleotides were found, is shown in
Figure 2 (line 1); the sequence interval was U920-A1046. Two
modi®ed nucleotides, C936 and C948, have been clearly and
reproducibly detected. Only two stops have been selected as
they are the only ones which do not have any detectable
counterparts in the control lines 2 and 3 (Fig. 2). The
differences in the modi®cation pattern from the previous
results (17) are probably due to the fact that here much lower
(sub-stoichiometric) amounts of Tc were used.